Q47. Retikulárna formácia mozgového kmeňa
Pojem neuroglia.
Neuroglia- sú to bunky, ktoré obklopujú neuróny a sú s nimi zahrnuté v centrálnom nervovom systéme a PNS. Počet gliových buniek je rádovo vyšší ako počet nervových buniek.
Funkcie neuroglie:
1.podporná – podporuje nervové bunky
2.izolačné - bráni prechodu nervových vzruchov z tela jedného neurónu do tela druhého
3.regulačné - podieľa sa na regulácii centrálneho nervového systému, najmä zabezpečuje prenos vzruchov správnym smerom
4.trofický – zúčastňuje sa na metabolických procesoch neurónov
5. regulačná – reguluje dráždivosť nervových buniek.
Membránový potenciál (alebo pokojový potenciál) je potenciálny rozdiel medzi vonkajším a vnútorným povrchom membrány v stave relatívneho fyziologického pokoja. Oddychový potenciál Vyvstáva v dôsledku dvoch dôvodov:
1) nerovnomerné rozloženie iónov na oboch stranách membrány;
2) selektívna permeabilita membrány pre ióny. V pokoji nie je membrána rovnaká priepustná pre rôzne ióny. Bunková membrána je priepustná pre ióny K, málo priepustná pre ióny Na a nepriepustná pre organické látky.
Vďaka týmto dvom faktorom sa vytvárajú podmienky pre pohyb iónov. Tento pohyb sa uskutočňuje bez spotreby energie pasívnym transportom - difúziou v dôsledku rozdielu v koncentrácii iónov. K ióny opúšťajú bunku a zvyšujú kladný náboj na vonkajšom povrchu membrány, Cl ióny pasívne prechádzajú do bunky, čo vedie k zvýšeniu kladného náboja na vonkajšom povrchu bunky. Ióny Na sa hromadia na vonkajšom povrchu membrány a zvyšujú jej kladný náboj. Organické zlúčeniny zostávajú vo vnútri bunky. V dôsledku tohto pohybu je vonkajší povrch membrány nabitý kladne a vnútorný povrch záporne. Vnútorný povrch membrány nemusí byť absolútne záporne nabitý, ale je vždy záporne nabitý vzhľadom na vonkajší. Tento stav bunkovej membrány sa nazýva stav polarizácie. Pohyb iónov pokračuje, kým sa potenciálny rozdiel cez membránu nevyrovná, t.j. nenastane elektrochemická rovnováha. Moment rovnováhy závisí od dvoch síl:
1) difúzne sily;
2) silu elektrostatická interakcia. Hodnota elektrochemickej rovnováhy:
1) udržiavanie iónovej asymetrie;
2) udržiavanie hodnoty membránového potenciálu na konštantnej úrovni.
Difúzna sila (rozdiel koncentrácie ióny) a sila elektrostatickej interakcie, preto sa membránový potenciál nazýva koncentračno-elektrochemický.
Elektrochemická rovnováha je nedostatočná na udržanie iónovej asymetrie. V klietke existujeďalším mechanizmom je sodíkovo-draslíková pumpa. Sodno-draselná pumpa - mechanizmus zabezpečenie aktívneho transportu iónov. Bunková membrána obsahuje systém nosičov, z ktorých každý viaže tri Na ióny, ktoré sú vnútri bunky a vynesie ich von. Zvonka sa nosič viaže na dva K ióny mimo bunky a prenáša ich do cytoplazmy. Energia sa získava z rozkladu ATP.
2) (Mechanizmus pokojového potenciálu)
Akčný potenciál je posun membránového potenciálu, vznikajúce v tkaniny pôsobením prahového a nadprahového podnetu, ktorý je sprevádzaný dobíjaním bunkovej membrány.
Pôsobením prahového alebo nadprahového podnetu sa mení priepustnosť bunková membrána pre ióny v rôznej miere. Pre ióny Na sa zvyšuje a gradient sa vyvíja pomaly. V dôsledku toho dochádza k pohybu iónov Na vo vnútri bunky, ióny Presťahovať sa z klietky, že vedie na dobitie bunkovej membrány. Vonkajší povrch membrány nesie záporný náboj, vnútorný povrch je kladný.
Komponenty akčného potenciálu:
1) lokálna odozva;
2) špičkový potenciál vysokého napätia (špička);
3) fluktuácie stopy.
Na ióny vstupujú do bunky jednoduchou difúziou bez spotreby energie. Dosiahnutie prahu silu membránový potenciál klesá na kritickú úroveň depolarizácie (asi 50 mV). Kritická úroveň depolarizácie je počet milivoltov, o ktorý by mal znižujú membránový potenciál tak, že nastáva lavínovitý priebeh iónov Na do bunky.
Špičkový potenciál vysokého napätia (špička).
Vrchol akčného potenciálu je konštantnou zložkou akčného potenciálu. Pozostáva z dvoch fáz:
1) vzostupná časť - fáza depolarizácie;
2) zostupná časť - fáza repolarizácie.
Lavínový tok iónov Na do bunky vedie k zmene potenciálu na bunkovej membráne. Čím viac iónov Na vstúpi do bunky, tým viac je membrána depolarizovaná, tým viac sa otvorí aktivačná brána. Vzhľad náboja s opačným znamienkom sa nazýva inverzia membránového potenciálu. Pohyb iónov Na vo vnútri článku pokračuje až do okamihu elektrochemickej rovnováhy vzhľadom na ión Na.Amplitúda akčného potenciálu nezávisí od sily podnetu, závisí od koncentrácie iónov Na a od stupňa priepustnosti membrán pre ióny Na. Zostupná fáza (fáza repolarizácie) vracia náboj membrány do pôvodného znamienka. Po dosiahnutí elektrochemickej rovnováhy pre ióny Na sa aktivačná brána deaktivuje, priepustnosť na ióny Na a zvyšuje sa priepustnosť pre ióny K. K úplnej obnove membránového potenciálu nedochádza.
V procese obnovy reakcie na bunkovej membráne sú zaznamenané stopové potenciály – pozitívne a negatívne.
3) (Zmena excitability pri prechode vlny vzrušenia)
S rozvojom akčného potenciálu dochádza k zmene dráždivosti tkaniva, pričom táto zmena prebieha vo fázach (obr. 2). Stav počiatočnej polarizácie membrány, ktorý sa odráža na pokojovom membránovom potenciáli, zodpovedá počiatočnému stavu jej excitability, a preto sú bunky na normálnej úrovni excitability. Počas obdobia pred spájkovaním sa zvyšuje excitabilita tkaniva, táto fáza excitability sa nazýva primárne povznesenie. Počas vývoja predspájkovania sa pokojový membránový potenciál blíži ku kritickej úrovni depolarizácie a na jej dosiahnutie stačí sila stimulu, ktorá je menšia ako prahová (podprahová).
V období vývoja hrotu (vrcholového potenciálu) dochádza k lavínovitému prúdeniu sodíkových iónov do bunky, v dôsledku čoho sa membrána dobíja a stráca schopnosť reagovať excitáciou na podnety aj nadprahovej sily. . Táto fáza excitability sa nazýva absolútna žiaruvzdornosť(absolútna nevzrušivosť). Vydrží až do konca dobitia membrány. Absolútna žiaruvzdornosť, teda úplná neexcitabilita membrány, vzniká v dôsledku skutočnosti, že sodíkové kanály sú na začiatku úplne otvorené a potom inaktivované.
Po ukončení fázy nabíjania membrány sa jej excitabilita postupne obnoví na pôvodnú úroveň - fáza relatívnej refraktérnosti. Pokračuje, kým sa náboj membrány neobnoví na hodnotu zodpovedajúcu kritickej úrovni depolarizácie. Keďže v tomto období ešte nebol obnovený pokojový membránový potenciál, dráždivosť tkaniva je znížená a nová excitácia môže vzniknúť len pôsobením nadprahového podnetu. Zníženie excitability vo fáze relatívnej refraktérnosti je spojené s čiastočnou inaktiváciou sodíkových kanálov a aktiváciou draslíkových kanálov.
Obdobie negatívneho stopového potenciálu zodpovedá zvýšenej úrovni excitability - fáza sekundárnej exaltácie. Keďže membránový potenciál je v tejto fáze bližšie ku kritickej úrovni depolarizácie, ale v porovnaní so stavom pokoja (počiatočná polarizácia) je prah podráždenia znížený, t.j. zvýšená excitabilita. V tejto fáze môže pôsobením podnetov podprahovej sily vzniknúť nové vzrušenie. Sodíkové kanály v tejto fáze nie sú úplne inaktivované. Počas obdobia rozvoja pozitívneho stopového potenciálu sa znižuje excitabilita tkaniva - fáza sekundárnej žiaruvzdornosti. Počas tejto fázy sa membránový potenciál zvyšuje (stav membránovej hyperpolarizácie), vzďaľuje sa od kritickej úrovne depolarizácie, stúpa prah podráždenia a nová excitácia môže vzniknúť len pôsobením podnetov s nadprahovou hodnotou. Hyperpolarizácia membrány sa vyvíja z troch dôvodov: po prvé, pokračujúce uvoľňovanie iónov draslíka; po druhé, prípadne otvorením kanálov pre chlór a vstupom týchto iónov do cytoplazmy bunky; po tretie, zvýšená práca sodíkovo-draselnej pumpy.
4) (Vedenie vzruchu pozdĺž nervových vlákien)
Mechanizmus šírenia vzruchu v rôznych nervových vláknach nie je rovnaký. Podľa moderných koncepcií sa šírenie vzruchu pozdĺž nervových vlákien uskutočňuje na základe iónových mechanizmov na generovanie akčného potenciálu.
S šírením excitácie pozdĺž nervového vlákna bez myelínu spôsobujú lokálne elektrické prúdy, ktoré vznikajú medzi jeho excitovanou časťou, negatívne nabitou a neexcitovanou, pozitívne nabitou, depolarizáciu membrány na kritickú úroveň, po ktorej nasleduje generovanie AP v najbližšom bode. neexcitovanej časti membrány. Tento proces sa mnohokrát opakuje. Po celej dĺžke nervového vlákna prebieha proces reprodukcie nového AP v každom bode membrány vlákna. Toto nazývate správanie sa vzrušenia a nepretržitý.
Prítomnosť plášťa s vysokým elektrickým odporom v myelínových vláknach, ako aj časti vlákien bez plášťa (zachytenia Ranviera) vytvárajú podmienky pre kvalitatívne nový typ vedenia vzruchu po myelinizovaných nervových vláknach. Miestne elektrické prúdy vznikajú medzi susednými Ranvierovými záchytmi, pretože excitovaná záchytná membrána sa stáva záporne nabitá vzhľadom na povrch susedného nevybudeného záchytného bodu. Tieto lokálne prúdy depolarizujú membránu neexcitovaného záchytu na kritickú úroveň a objaví sa v nej AP (obr. 4). V dôsledku toho sa zdá, že vzrušenie „preskakuje“ cez oblasti nervového vlákna pokryté myelínom z jedného záberu do druhého. Takýto mechanizmus šírenia vzruchu sa nazýva soľný alebo kŕčovitý. Rýchlosť tohto spôsobu vedenia budenia je oveľa vyššia a je ekonomickejšia v porovnaní s kontinuálnym vedením budenia, pretože do stavu aktivity nie je zapojená celá membrána, ale iba jej malé časti v oblasti odpočúvania.
Ryža. 4. Schéma šírenia vzruchu v bezmyelínových (a) a myelínových (b) nervových vláknach.
"Preskočenie" akčného potenciálu cez úsek medzi odposluchmi je možné, pretože amplitúda AP je 5-6 krát vyššia ako prahová hodnota potrebná na vybudenie susedného odposluchu. PD môže „preskočiť“ nielen cez jednu, ale aj cez dve medzizáchytné medzery. Tento jav možno pozorovať pri znížení excitability susedného záchytu pod vplyvom akejkoľvek farmakologickej látky, napríklad novokaínu, kokaínu atď.
Nervové vlákna majú labilita- schopnosť reprodukovať určitý počet cyklov vzruchu za jednotku času v súlade s rytmom pôsobiacich podnetov. Mierou lability je maximálny počet excitačných cyklov, ktoré môže nervové vlákno reprodukovať za jednotku času bez transformácie stimulačného rytmu. Labilita je určená trvaním vrcholu akčného potenciálu, teda fázou absolútnej refraktérnosti. Keďže trvanie absolútnej refrakcie v hrotovom potenciáli nervového vlákna je najkratšie, jeho labilita je najvyššia. Nervové vlákno je schopné reprodukovať až 1000 impulzov za sekundu.
N.E. Vvedensky zistil, že ak je odkrytá časť nervu zmeny(t. j. vystavenie škodlivému činidlu) napríklad otravou alebo poškodením, potom sa labilita takéhoto miesta výrazne zníži. K obnove počiatočného stavu nervového vlákna po každom akčnom potenciáli v poškodenej oblasti dochádza pomaly. Keď je toto miesto vystavené častým podnetom, nie je schopné reprodukovať daný rytmus stimulácie, a preto je vedenie impulzov zablokované. Tento stav zníženej lability pomenoval N.E. Vvedensky parabióza. Vo vývoji stavu parabiózy možno zaznamenať tri po sebe nasledujúce fázy: vyrovnávaciu, paradoxnú, inhibičnú.
V vyrovnávacia fáza dochádza k vyrovnaniu veľkosti odozvy na časté a zriedkavé podnety. Za normálnych podmienok fungovania nervového vlákna sa veľkosť odozvy ním inervovaných svalových vlákien riadi zákonom sily: odozva na vzácne podnety je menšia a na časté podnety väčšia. Pôsobením parabiotického činidla a so zriedkavým rytmom podráždenia (napríklad 25 Hz) sú všetky excitačné impulzy vedené cez parabiotické miesto, pretože excitabilita po predchádzajúcom impulze má čas na zotavenie. Pri vysokom rytme stimulácie (100 Hz) môžu následné impulzy doraziť v čase, keď je nervové vlákno ešte v stave relatívnej refraktérnosti spôsobenej predchádzajúcim akčným potenciálom. Preto niektoré impulzy nie sú vedené. Ak sa vykoná len každá štvrtá excitácia (to znamená 25 impulzov zo 100), potom sa amplitúda odozvy stane rovnakou ako pri zriedkavých podnetoch (25 Hz) - dôjde k vyrovnaniu odozvy.
V paradox-fáza dochádza k ďalšiemu poklesu lability. Súčasne dochádza k reakcii na zriedkavé a časté podnety, ale oveľa menej na časté podnety, pretože časté podnety ďalej znižujú labilitu a predlžujú fázu absolútnej refraktérnosti. Následne dochádza k paradoxu – odozva na vzácne podnety je väčšia ako na časté.
V fáza brzdenia labilita klesá do takej miery, že zriedkavé aj časté podnety nevyvolávajú reakciu. V tomto prípade je membrána nervového vlákna depolarizovaná a neprechádza do štádia repolarizácie, to znamená, že jej pôvodný stav nie je obnovený.
Fenomén parabiózy je základom lokálnej anestézie vyvolanej liekmi. Účinok anestetík je tiež spojený so znížením lability a porušením mechanizmu vedenia vzruchu pozdĺž nervových vlákien.
Parabióza je reverzibilný jav. Ak parabiotická látka pôsobí krátkodobo, tak po ukončení jej pôsobenia nerv opúšťa stav parabiózy cez rovnaké fázy, ale v opačnom slede.
Mechanizmus vývoja parabiotického stavu je nasledujúci. Keď parabiotický faktor pôsobí na nervové vlákno, schopnosť membrány zvyšovať priepustnosť sodíka v reakcii na podráždenie je narušená. V oblasti zmeny sa inaktivácia sodíkových kanálov spôsobená škodlivým činidlom kombinuje s inaktiváciou spôsobenou nervovým impulzom a excitabilita sa zníži natoľko, že sa blokuje vedenie ďalšieho impulzu.
5) (Synapsie, ich typy, štrukturálne znaky)
Fyziológia synapsií.
V CNS sú nervové bunky navzájom spojené prostredníctvom synapsií. Synapse Je štrukturálne funkčný útvar, ktorý zabezpečuje prenos vzruchu alebo inhibície z nervového vlákna do inervovanej bunky.
Synapsie o lokalizácii sa delia na centrálne (nachádzajú sa v rámci centrálneho nervového systému, ako aj v gangliách autonómneho nervového systému) a periférne (umiestnené mimo centrálneho nervového systému, zabezpečujú komunikáciu s bunkami inervovaného tkaniva).
Funkčne synapsie sa delia na vzrušujúce, pri ktorej v dôsledku depolarizácie postsynaptickej membrány vzniká excitačný postsynaptický potenciál a brzda, v ktorých presynaptických zakončeniach sa uvoľňuje mediátor, hyperpolarizujúci postsynaptickú membránu a spôsobujúci vznik inhibičného postsynaptického potenciálu.
Prevodovým mechanizmom synapsie sa delia na chemické a elektrické. Chemické synapsie prenášajú vzrušenie alebo inhibíciu vďaka špeciálnym látkam - mediátorom. Záleží na od typu mediátora chemické synapsie sa delia na:
1.cholinergikum (mediátor - acetylcholín)
2.adrenergné (mediátory - adrenalín, norepinefrín)
Anatomická klasifikácia synapsie sa delia na neurosekrečné, neuromuskulárne a interneuronálne.
Synapse pozostáva z troch hlavných komponentov:
1.presynaptická membrána
2.Postsynaptická membrána
3.synaptická štrbina
Presynaptická membrána je koniec procesu nervovej bunky. Vo vnútri prílohy, v bezprostrednej blízkosti membrány, dochádza k nahromadeniu vezikúl (granúl) obsahujúcich jeden alebo iný mediátor. Bubliny sú v neustálom pohybe.
Postsynaptická membrána je súčasťou bunkovej membrány inervovaného tkaniva. Postsynaptická membrána na rozdiel od presynaptickej membrány má proteínové chemoreceptory na biologicky aktívne (mediátory, hormóny), liečivé a toxické látky. Dôležitou vlastnosťou postsynaptických membránových receptorov je ich chemická špecifickosť, t.j. schopnosť vstúpiť do biochemickej interakcie len s určitým typom mediátora.
Synaptická štrbina je priestor medzi pre- a postsynaptickou membránou, vyplnený kvapalinou podobnou zložením ako krvná plazma. Prostredníctvom nej mediátor pomaly difunduje z presynaptickej membrány do postsynaptickej membrány.
Štrukturálne znaky neuromuskulárnej synapsie určujú jej fyziologické vlastnosti.
1. Jednostranné vedenie vzruchu (z pre- k postsynaptickej membráne), v dôsledku prítomnosti receptorov citlivých na mediátor iba v postsynaptickej membráne.
2. Synaptické oneskorenie vo vedení vzruchu (čas medzi príchodom impulzu na presynaptickom konci a nástupom postsynaptickej odpovede), spojené s nízkou rýchlosťou difúzie vysielača do synaptickej štrbiny v porovnaní s rýchlosťou impulz cez nervové vlákno.
3. Nízka labilita a vysoká únava synapsie v dôsledku času šírenia predchádzajúceho impulzu a prítomnosti obdobia absolútnej refraktérnosti v ňom.
4. Vysoká selektívna citlivosť synapsie na chemikálie vďaka špecifickosti chemoreceptorov postsynaptických membrán.
Etapy synaptického prenosu.
1. Syntéza mediátora... V cytoplazme neurónov a nervových zakončení sa syntetizujú chemické mediátory - biologicky aktívne látky. Neustále sa syntetizujú a ukladajú v synaptických vezikulách nervových zakončení.
2. Sekrécia vysielača... Uvoľňovanie neurotransmiteru zo synaptických vezikúl je kvantovej povahy. V pokoji je nevýznamný, ale pod vplyvom nervového impulzu sa prudko zvyšuje.
3. Interakcia mediátora s receptormi postsynaptickej membrány... Táto interakcia spočíva v selektívnej zmene permeability iónovo selektívnych kanálov efektorovej bunky v oblasti aktívnych centier väzby na mediátor. Interakcia mediátora s jeho receptormi môže spôsobiť excitáciu alebo inhibíciu neurónu, kontrakciu svalovej bunky, tvorbu a uvoľňovanie hormónov sekrečnými bunkami. V prípade zvýšenia permeability sodíkových a vápnikových kanálov sa zvyšuje prietok Na a Ca do bunky, nasleduje depolarizácia membrány, vznik PD a ďalší prenos nervového vzruchu. Takéto synapsie sa nazývajú excitačné. Ak sa zvyšuje priepustnosť draslíkových kanálov a kanálov pre chlór, dochádza k nadmernému uvoľňovaniu K z bunky so súčasnou difúziou Cl do nej, čo vedie k hyperpolarizácii membrány, zníženiu jej excitability a rozvoju inhibičných postsynaptických potenciálov. . Prenos nervových impulzov sa sťaží alebo sa úplne zastaví. Takéto synapsie sa nazývajú inhibičné synapsie.
Receptory, ktoré interagujú s ACh, sa nazývajú cholinergné receptory. Funkčne sa delia do dvoch skupín: M - a H-cholinergné receptory. V synapsiách kostrových svalov sú prítomné iba H-cholinergné receptory, zatiaľ čo vo svaloch vnútorných orgánov sú prítomné hlavne M-cholinergné receptory.
Receptory, ktoré interagujú s HA, sa nazývajú adrenergné receptory. Funkčne sa delia na alfa a beta adrenergné receptory. V postsynaptickej membráne buniek hladkého svalstva vnútorných orgánov a krvných ciev často koexistujú oba typy adrenergných receptorov. Pôsobenie NA je depolarizačné, ak interaguje s alfa-adrenergnými receptormi (kontrakcia svalovej membrány stien krvných ciev alebo čriev), alebo inhibičné - keď interaguje s beta-adrenergnými receptormi (ich relaxácia).
4. Deaktivácia vysielača... Inaktivácia (úplná strata aktivity) mediátora je nevyhnutná pre repolarizáciu postsynaptickej membrány a obnovenie počiatočnej hladiny MP. Najdôležitejšou cestou inaktivácie je hydrolytická degradácia inhibítormi. Pre ACh je inhibítorom cholínesteráza, pre HA a adrenalín - monoaminooxidáza a katecholoxymetyltransferáza.
Ďalším spôsobom odstránenia mediátora zo synaptickej štrbiny je „spätné vychytávanie“ jeho presynaptickými zakončeniami (pinocytóza) a reverzný axonálny transport, obzvlášť výrazný u katecholamínov.
Koordinačná činnosť centrálneho nervového systému je založená na interakcii procesov excitácie a inhibície.
Vzrušenie Je aktívny proces, ktorý je reakciou tkaniva na podráždenie a je charakterizovaný zvýšením funkcií tkaniva.
Brzdenie Je aktívny proces, ktorý je reakciou tkaniva na podráždenie a je charakterizovaný znížením funkcií tkaniva.
Primárna inhibícia v centrálnom nervovom systéme nastáva v dôsledku inhibičných neurónov. Ide o špeciálny typ interkalárnych neurónov, ktoré pri prenose impulzu uvoľňujú inhibičný neurotransmiter. Existujú 2 typy primárnej inhibície: postsynaptická a presynaptická.
Postsynaptická inhibícia nastane, ak axón inhibičného neurónu vytvorí synapsiu s telom neurónu a uvoľnením vysielača spôsobí hyperpolarizáciu bunkovej membrány, čo inhibuje aktivitu bunky.
Presynaptická inhibícia nastáva, keď axón inhibičného neurónu tvorí synapsiu s axónom excitačného neurónu, čo interferuje s vedením impulzu.
6) (Miecha, jej funkcie, účasť na regulácii svalového tonusu)
Miecha vykonáva reflexné a vodivé funkcie. Prvú zabezpečujú jej nervové centrá, druhú dráhy.
Má segmentovú štruktúru. Rozdelenie na segmenty je navyše funkčné. Každý segment tvorí predné a zadné korene. Zadné sú citlivé, t.j. aferentný, predný motorický, eferentný. Tento vzorec sa nazýva Bella-Magendieho zákon. Korene každého segmentu inervujú 3 telesné metaméry, ale v dôsledku prekrývania je každá metaméra inervovaná tromi segmentmi. Preto, keď sú ovplyvnené predné korene jedného segmentu, motorická aktivita zodpovedajúceho metaméru je iba oslabená.
Morfologicky tvoria telá neurónov miechy jej šedú hmotu. Funkčne sú všetky jeho neuróny rozdelené na motoneuróny, interkalárne, neuróny sympatického a parasympatického oddelenia autonómneho nervového systému. Motorické neuróny sa v závislosti od ich funkčného významu delia na alfa a gama motorické neuróny. Vlákna aferentných dráh smerujú do a-motoneurónov, ktoré vychádzajú z intrafuzálnych, t.j. receptorové svalové bunky. Telá a-motorických neurónov sa nachádzajú v predných rohoch miechy a ich axóny inervujú kostrové svaly. Gama motoneuróny regulujú napätie svalového vretena t.j. intrafúzne vlákna. Podieľajú sa teda na regulácii kontrakcií kostrového svalstva. Preto, keď sú predné korene prerezané, svalový tonus zmizne.
Interneuróny zabezpečujú komunikáciu medzi centrami miechy a priľahlými časťami centrálneho nervového systému.
Neuróny sympatickej časti autonómneho nervového systému sa nachádzajú v bočných rohoch hrudných segmentov a parasympatikus v sakrálnej časti.
Vodivou funkciou je zabezpečiť prepojenie periférnych receptorov, centier miechy s nadložnými časťami centrálneho nervového systému, ako aj jeho nervových centier navzájom. Vykonáva sa vodivými dráhami. Všetky dráhy miechy sú rozdelené na vlastné alebo propriospinálne, vzostupné a zostupné. Propriospinálne dráhy spájajú nervové centrá rôznych segmentov miechy. Ich funkciou je koordinovať svalový tonus, pohyby rôznych metamérov trupu.
Vzostupné cesty zahŕňajú niekoľko ciest. Gaullove a Burdachove zväzky vedú nervové impulzy z proprioceptorov svalov a šliach do zodpovedajúcich jadier predĺženej miechy a potom do talamu a somatosenzorických oblastí kôry. Vďaka týmto dráham sa posudzuje a koriguje držanie tela. Zväzky Govers a Fleksig prenášajú vzruch z proprioceptorov, mechanoreceptorov kože do mozočku. Vďaka tomu je zabezpečené vnímanie a nevedomá koordinácia držania tela. Spinothalamické dráhy vedú signály od bolesti, teploty, hmatových receptorov kože do talamu a potom do somatosenzorických zón kôry. Zabezpečujú vnímanie zodpovedajúcich signálov a vytváranie citlivosti.
Zostupné cesty sú tiež tvorené niekoľkými cestami. Kortikospinálne dráhy idú z pyramídových a extrapyramídových neurónov kôry do a-motorických neurónov miechy. Vďaka nim sa vykonáva regulácia dobrovoľných pohybov. Rubrospinálna dráha vedie signály z červeného jadra stredného mozgu do a-motorických neurónov flexorových svalov. Vestibulospinálna dráha prenáša signály z vestibulárnych jadier medulla oblongata, predovšetkým Deitersovho jadra, do a-motorických neurónov extenzorových svalov. Vďaka týmto dvom dráham je tonus zodpovedajúcich svalov regulovaný zmenami polohy tela.
Všetky reflexy miechy sa delia na somatické, t.j. motorické a vegetatívne. Somatické reflexy sa delia na šľachové alebo myotické a kožné. Šľachové reflexy vznikajú pri mechanickom podráždení svalov a šliach. Ich mierne natiahnutie vedie k excitácii šľachových receptorov a a-motoneurónov miechy. Výsledkom je kontrakcia svalov, predovšetkým extenzorov. Šľachové reflexy zahŕňajú koleno, Achillovu, lakeť, zápästie atď., vznikajúce mechanickým podráždením príslušných šliach. Napríklad koleno je najjednoduchšie monosynaptické, pretože v jeho centrálnej časti je iba jedna synapsia. Kožné reflexy sú spôsobené podráždením kožných receptorov, ale prejavujú sa motorickými reakciami. Sú plantárne a brušné (vysvetlenie). Miechové nervové centrá sú pod kontrolou nadložných. Preto po úseku medzi predĺženou miechou a miechou nastáva miechový šok a výrazne sa zníži tonus všetkých svalov.
Autonómne reflexy miechy sa delia na sympatické a parasympatické. Obe sa prejavujú reakciou vnútorných orgánov na podráždenie receptorov kože, vnútorných orgánov, svalov. Vegetatívne neuróny miechy tvoria spodné centrá pre reguláciu cievneho tonusu, srdcovej činnosti, priesvitu priedušiek, potenia, močenia, defekácie, erekcie, ejakulácie atď.
7) (Predĺžená dreň a mostík, ich funkcie, účasť na regulácii svalového tonusu)
Medulla
Vlastnosti funkčnej organizácie. Medulla oblongata (medullaoblongata) u ľudí má dĺžku asi 25 mm. Ide o predĺženie miechy. Štrukturálne, pokiaľ ide o rozmanitosť a štruktúru jadier, je predĺžená miecha zložitejšia ako miecha. Na rozdiel od miechy nemá metamérnu, opakovateľnú štruktúru, sivá hmota v nej nie je umiestnená v strede, ale v jadrách smerom k periférii.
V predĺženej mieche sú olivy spojené s miechou, extrapyramídovým systémom a mozočkom – ide o tenké a klinovité jadrá proprioceptívnej citlivosti (jadrá Gaulle a Burdach). Sú tu aj priesečníky klesajúcich pyramídových dráh a vzostupných dráh tvorených tenkými a klinovitými zväzkami (Gaulle a Burdach), retikulárny útvar.
Medulla oblongata sa vďaka svojim jadrovým formáciám a retikulárnej formácii podieľa na realizácii autonómnych, somatických, chuťových, sluchových, vestibulárnych reflexov. Charakteristickým znakom medulla oblongata je, že jej jadrá, ktoré sú excitované postupne, poskytujú komplexné reflexy, ktoré si vyžadujú sekvenčnú aktiváciu rôznych svalových skupín, čo sa pozoruje napríklad pri prehĺtaní.
V medulla oblongata sa nachádzajú jadrá nasledujúcich hlavových nervov:
pár VIII hlavových nervov - vestibulárny kochleárny nerv pozostáva z kochleárnej a vestibulárnej časti. Kochleárne jadro leží v medulla oblongata;
pár IX - glossofaryngeálny nerv (položka glossopharyngeus); jeho jadro tvoria 3 časti – motorická, senzitívna a vegetatívna. Motorická časť sa podieľa na inervácii svalov hltana a ústnej dutiny, senzitívna časť prijíma informácie z chuťových receptorov zadnej tretiny jazyka; vegetatívny inervuje slinné žľazy;
pár X - blúdivý nerv (n.vagus) má 3 jadrá: vegetatívne inervuje hrtan, pažerák, srdce, žalúdok, črevá, tráviace žľazy; citlivý prijíma informácie z receptorov alveol pľúc a iných vnútorných orgánov a motorický (tzv. vzájomný) zabezpečuje sekvenciu kontrakcie svalov hltana a hrtana pri prehĺtaní;
pár XI - prídavný nerv (n.accessorius); jeho jadro je čiastočne umiestnené v medulla oblongata;
pár XII - nerv hypoglossus (n.hypoglossus) je motorický nerv jazyka, jeho jadro sa väčšinou nachádza v medulla oblongata.
Senzorické funkcie. Medulla oblongata reguluje množstvo zmyslových funkcií: príjem kožnej citlivosti tváre - v senzorickom jadre trojklaného nervu; primárna analýza príjmu chuti - v jadre glosofaryngeálneho nervu; príjem sluchových podnetov - v jadre kochleárneho nervu; príjem vestibulárnych podnetov - v hornom vestibulárnom jadre. V zadných horných častiach medulla oblongata sú dráhy kožnej, hlbokej, viscerálnej citlivosti, z ktorých niektoré sú tu prepojené na druhý neurón (tenké a klinovité jadrá). Na úrovni medulla oblongata vyššie uvedené senzorické funkcie implementujú primárnu analýzu sily a kvality stimulácie, následne sa spracované informácie prenesú do subkortikálnych štruktúr, aby sa určil biologický význam tejto stimulácie.
Vedenie funkcií. Všetky vzostupné a zostupné cesty miechy prechádzajú cez medulla oblongata: spinálny talamický, kortikospinálny, rubrospinálny. Pochádza z vestibulospinálneho, olivospinálneho a retikulospinálneho traktu, ktoré zabezpečujú tonus a koordináciu svalových reakcií. V medulla oblongata končia dráhy z mozgovej kôry - kortex-retikulárne dráhy. Tu končia vzostupné dráhy proprioceptívnej citlivosti z miechy: tenké a klinovité. Takéto formácie mozgu ako most, stredný mozog, mozoček, talamus, hypotalamus a mozgová kôra majú obojsmerné spojenie s medulla oblongata. Prítomnosť týchto spojení naznačuje účasť medulla oblongata na regulácii tonusu kostrového svalstva, autonómnych a vyšších integračných funkciách a analýze senzorických stimulov.
Reflexné funkcie. Početné reflexy medulla oblongata sú rozdelené na vitálne a neživotné, avšak tento koncept je skôr svojvoľný. Respiračné a vazomotorické centrá medulla oblongata možno pripísať životne dôležitým centrám, pretože v nich je uzavretých množstvo srdcových a respiračných reflexov.
Medulla oblongata organizuje a realizuje množstvo ochranných reflexov: vracanie, kýchanie, kašeľ, slzenie, zatváranie viečok. Tieto reflexy sa realizujú vďaka tomu, že informácie o podráždení receptorov sliznice oka, ústnej dutiny, hrtana, nosohltanu cez citlivé vetvy trigeminálneho a glosofaryngeálneho nervu vstupujú do jadier predĺženej miechy, odtiaľ do príkaz ide do motorických jadier trigeminálneho, vagusového, tvárového, glosofaryngeálneho, prídavného alebo hypoglossálneho nervu, v dôsledku čoho sa realizuje jeden alebo iný ochranný reflex. Rovnakým spôsobom sa v dôsledku postupného zaraďovania svalových skupín hlavy, krku, hrudníka a bránice organizujú reflexy stravovacieho správania: sanie, žuvanie, prehĺtanie.
Okrem toho medulla oblongata organizuje reflexy udržiavania polohy. Tieto reflexy sa tvoria v dôsledku aferentácie z receptorov na vestibule kochley a polkruhových kanálov do horného vestibulárneho jadra; odtiaľto sa spracované informácie na posúdenie potreby zmeny držania tela posielajú do laterálnych a mediálnych vestibulárnych jadier. Tieto jadrá sa podieľajú na určovaní toho, ktoré svalové systémy, segmenty miechy by sa mali podieľať na zmene držania tela, preto signál ide z neurónov mediálneho a laterálneho jadra pozdĺž vestibulospinálnej dráhy do predných rohov zodpovedajúcich segmentov. miechy, inervujúce svaly, ktorých účasť na zmene držania tela v danom momente je nevyhnutná.
Zmena držania tela sa uskutočňuje v dôsledku statických a statokinetických reflexov. Statické reflexy regulujú tonus kostrového svalstva, aby sa udržala určitá poloha tela. Statokinetické reflexy medulla oblongata poskytujú redistribúciu tonusu svalov trupu, aby organizovali pózu zodpovedajúcu momentu priamočiareho alebo rotačného pohybu.
Väčšina autonómnych reflexov medulla oblongata sa realizuje cez jadrá vagusového nervu v nej umiestnené, ktoré dostávajú informácie o stave činnosti srdca, ciev, tráviaceho traktu, pľúc, tráviacich žliaz atď. tieto informácie, jadrá organizujú motorické a sekrečné reakcie týchto orgánov.
Excitácia jadier blúdivého nervu spôsobuje zvýšenie kontrakcie hladkých svalov žalúdka, čriev, žlčníka a zároveň uvoľnenie zvieračov týchto orgánov. Súčasne sa práca srdca spomaľuje a oslabuje, lúmen priedušiek sa zužuje.
Činnosť jadier blúdivého nervu sa prejavuje aj zvýšením sekrécie bronchiálnych, žalúdočných, črevných žliaz, pri excitácii pankreasu, sekrečných buniek pečene.
V medulla oblongata je lokalizované centrum slinenia, ktorého parasympatická časť zabezpečuje zvýšenie všeobecnej sekrécie a sympatická časť sekrécie bielkovín slinných žliaz.
Respiračné a vazomotorické centrá sa nachádzajú v štruktúre retikulárnej formácie medulla oblongata. Zvláštnosťou týchto centier je, že ich neuróny sú schopné byť excitované reflexne a pod vplyvom chemických podnetov.
Dýchacie centrum je lokalizované v strednej časti retikulárnej formácie každej symetrickej polovice medulla oblongata a je rozdelené na dve časti, inhaláciu a výdych.
V retikulárnej formácii medulla oblongata je zastúpené ďalšie životne dôležité centrum - vazomotorické centrum (regulácia cievneho tonusu). Funguje v spojení s nadložnými štruktúrami mozgu a predovšetkým s hypotalamom. Excitácia vazomotorického centra vždy mení rytmus dýchania, tonus priedušiek, svalov čriev, močového mechúra, ciliárneho svalu atď. Je to spôsobené tým, že retikulárna formácia predĺženej drene má synaptické spojenie hypotalamu a iných centier.
V stredných úsekoch retikulárnej formácie sa nachádzajú neuróny tvoriace retikulospinálnu dráhu, ktorá pôsobí inhibične na motorické neuróny miechy. V spodnej časti IV komory sa nachádzajú neuróny „modrej škvrny“. Ich mediátorom je norepinefrín. Tieto neuróny spôsobujú aktiváciu retikulospinálnej dráhy počas REM spánku, čo vedie k inhibícii miechových reflexov a zníženiu svalového tonusu.
Príznaky poškodenia. Poškodenie ľavej alebo pravej polovice medulla oblongata nad priesečníkom vzostupných dráh proprioceptívnej citlivosti spôsobuje poruchy citlivosti a práce svalov tváre a hlavy na strane poranenia. Zároveň na opačnej strane oproti strane poranenia dochádza k poruchám citlivosti kože a motorickej obrne trupu a končatín. Je to spôsobené tým, že vzostupné a zostupné dráhy z miechy a do miechy sa pretínajú a jadrá hlavových nervov inervujú svoju polovicu hlavy, čiže hlavové nervy sa nepretínajú.
Most
Mostík (ponscerebri, ponsVarolii) sa nachádza nad predĺženou miechou a vykonáva senzorické, vodivé, motorické, integračné reflexné funkcie.
Mostík zahŕňa jadrá tvárového nervu, trojklaného nervu, abducens, vestibulárny a kochleárny nerv (vestibulárne a kochleárne jadrá), jadrá vestibulárneho kochleárneho nervu (vestibulárny nerv): laterálny (Deiters) a horný (ankylozujúca spondylitída). Retikulárna formácia pons úzko súvisí s retikulárnou formáciou stredného mozgu a medulla oblongata.
Dôležitou štruktúrou mostíka je stredná stopka cerebellum. Práve ona zabezpečuje funkčné kompenzačné a morfologické spojenia mozgovej kôry s cerebelárnymi hemisférami.
Senzorické funkcie ponsu zabezpečujú jadrá vestibulárneho kochleárneho a trojklaného nervu. Kochleárna časť vestibulárneho kochleárneho nervu končí v mozgu v kochleárnych jadrách; vestibulárna časť vestibulárneho kochleárneho nervu - v trojuholníkovom jadre, Deitersovom jadre, ankylozujúcej spondylitis nucleus. Tu prebieha primárna analýza vestibulárnych stimulov ich sily a smeru.
Senzorické jadro trojklaného nervu prijíma signály z receptorov v koži tváre, prednej časti hlavy, slizniciach nosa a úst, zubov a spojoviek očnej buľvy. Lícny nerv (n. Facialis) inervuje všetky tvárové svaly tváre. Nerv abducens (n. Abducens) inervuje priamy laterálny sval, ktorý abdukuje očnú buľvu smerom von.
Motorická časť jadra trojklaného nervu (položka trigeminus) inervuje žuvacie svaly, sval, ktorý ťahá ušný bubienok, a sval, ktorý ťahá palatínový záves.
Vodivá funkcia mostíka. Opatrené pozdĺžne a priečne usporiadanými vláknami. Priečne umiestnené vlákna tvoria hornú a spodnú vrstvu a medzi nimi sú pyramídové dráhy vybiehajúce z mozgovej kôry. Medzi priečnymi vláknami sa nachádzajú nervové zhluky – jadrá mostíka. Z ich neurónov začínajú priečne vlákna, ktoré idú na opačnú stranu mosta, tvoria stredný pedikel mozočka a končia v jeho kôre.
V kryte mostíka sa nachádzajú pozdĺžne prebiehajúce zväzky vlákien mediálnej slučky (lemniscusmedialis). Prechádzajú nimi priečne vlákna lichobežníkového telesa (corpustrapezoideum), čo sú axóny kochleárnej časti vestibulárneho kochleárneho nervu opačnej strany, ktoré končia v jadre olivy nadradenej (olivasuperior). Z tohto jadra vychádzajú dráhy laterálnej slučky (lemniscus lateralis), ktoré smerujú do zadného štvoruholníka stredného mozgu a do mediálnych genikulárnych telies diencefala.
Predné a zadné jadrá lichobežníkového tela a laterálna slučka sú lokalizované v tektu. Tieto jadrá spolu s hornou olivou poskytujú primárnu analýzu informácií z orgánu sluchu a potom prenášajú informácie do zadných tuberkulov štvorcov.
Tektum obsahuje aj dlhé mediálne a tektospinálne dráhy.
Vlastné neuróny mostonosnej štruktúry tvoria jej retikulárnu formáciu, jadro tvárových, abdukčných nervov, motorickú časť jadra a stredné senzorické jadro trigeminálneho nervu.
Retikulárna formácia mosta je pokračovaním retikulárnej formácie medulla oblongata a začiatkom toho istého systému stredného mozgu. Axóny neurónov retikulárnej formácie pons idú do mozočku, do miechy (retikulospinálna dráha). Ten aktivuje neuróny miechy.
Retikulárna formácia mostíka ovplyvňuje mozgovú kôru, čo spôsobuje jej prebudenie alebo spánok. V retikulárnej formácii mostíka existujú dve skupiny jadier, ktoré patria do spoločného dýchacieho centra. Jedno centrum aktivuje inspiračné centrum medulla oblongata, druhé aktivuje výdychové centrum. Neuróny dýchacieho centra umiestnené v mostíku prispôsobujú prácu dýchacích buniek medulla oblongata v súlade s meniacim sa stavom tela.
8) (Stredný mozog, jeho funkcie, účasť na regulácii svalového tonusu)
Morfofunkčná organizácia. Stredný mozog (mezencefalón) predstavuje štvornásobok a nohy mozgu. Najväčšími jadrami stredného mozgu sú červené jadro, čierna substantia a jadrá hlavových (okulomotorických a blokových) nervov, ako aj jadro retikulárnej formácie.
Senzorické funkcie. Realizujú sa vďaka prijímaniu vizuálnych a sluchových informácií v ňom.
Vodivá funkcia. Spočíva v tom, že cez ňu prechádzajú všetky vzostupné dráhy do nadložného talamu (mediálna slučka, spinotalamická dráha), veľkého mozgu a mozočku. Zostupné cesty idú cez stredný mozog do predĺženej miechy a miechy. Ide o pyramídovú dráhu, vlákna kortikálneho mostíka, rubroretikulospinálnu dráhu.
Funkcia motora. Realizuje sa vďaka jadru blokového nervu (n. Trochlearis), jadrám okohybného nervu (it. oculomotorius), červenému jadru (nucleus ruber), čiernej hmote (substantianigra).
Červené jadrá sú umiestnené v hornej časti mozgových stopiek. Sú spojené s mozgovou kôrou (dráhy zostupujúce z kôry), subkortikálnymi jadrami, mozočkom, miechou (červená miechová dráha). Bazálne gangliá mozgu, cerebellum majú svoje konce v červených jadrách. Porušenie spojení červených jadier s retikulárnou formáciou medulla oblongata vedie k decerebračnej rigidite. Tento stav je charakterizovaný silným napätím extenzorových svalov končatín, krku, chrbta. Hlavnou príčinou decerebračnej rigidity je výrazný aktivačný účinok laterálneho vestibulárneho jadra (Deiters nucleus) na extenzorové motorické neuróny. Tento vplyv je maximálny pri absencii inhibičných vplyvov červeného jadra a nadložných štruktúr, ako aj cerebellum. Keď je mozog transekovaný pod jadrom laterálneho vestibulárneho nervu, decerebrálna rigidita zmizne.
Červené jadrá, ktoré prijímajú informácie z motorickej zóny mozgovej kôry, subkortikálnych jadier a mozočku o nadchádzajúcom pohybe a stave pohybového aparátu, vysielajú korekčné impulzy do motorických neurónov miechy pozdĺž rubrospinálneho traktu a tým regulujú sval tón, pripravuje jeho úroveň pre načrtnuté dobrovoľné hnutie ...
Ďalšie funkčne dôležité jadro stredného mozgu - substantia nigra - sa nachádza v nohách mozgu, reguluje úkony žuvania, prehĺtania (ich postupnosť), zabezpečuje presné pohyby prstov ruky napríklad pri písaní. Neuróny tohto jadra sú schopné syntetizovať mediátor dopamín, ktorý je dodávaný axonálnym transportom do bazálnych ganglií mozgu. Porážka substantia nigra vedie k porušeniu plastového tonusu svalov. Jemnú reguláciu plastického tónu pri hre na husle, písaní, vykonávaní grafických prác zabezpečuje čierna hmota. Zároveň pri dlhodobom udržiavaní určitého postoja dochádza vo svaloch k plastickým zmenám v dôsledku zmien ich koloidných vlastností, čo zabezpečuje najnižšiu spotrebu energie. Reguláciu tohto procesu vykonávajú bunky substantia nigra.
Neuróny jadier okulomotorických a blokových nervov regulujú pohyb oka hore, dole, von, k nosu a dole ku kútiku nosa. Neuróny prídavného jadra okulomotorického nervu (Jakubovičovo jadro) regulujú lúmen zrenice a zakrivenie šošovky.
Reflexné funkcie. Funkčne nezávislé štruktúry stredného mozgu sú kopčeky štvorice. Horné sú primárne subkortikálne centrá vizuálneho analyzátora (spolu s laterálnymi genikulárnymi telami diencefala), dolné sú sluchové (spolu so strednými genikulárnymi telami diencefala). Sú primárnym prepínaním vizuálnych a sluchových informácií. Z tuberkulov štvorky idú axóny ich neurónov do retikulárnej formácie kmeňa, motorických neurónov miechy. Štvornásobné neuróny môžu byť polymodálne a detektorové. V druhom prípade reagujú len na jeden príznak podráždenia, napríklad na zmenu svetla a tmy, smer pohybu svetelného zdroja atď. Aktivácia stredného mozgu v týchto prípadoch cez hypotalamus vedie k zvýšenému svalovému tonusu, zvýšeniu srdcovej frekvencie; existuje príprava na vyhýbanie sa, na obrannú reakciu.
Štvorka organizuje orientačné zrakové a sluchové reflexy.
U ľudí je štvornásobný reflex sentinelovým reflexom. V prípadoch zvýšenej vzrušivosti štvorčiat pri náhlej zvukovej alebo svetelnej stimulácii sa človek začne trhať, niekedy vyskočí na nohy, kričí, čo najrýchlejšie sa vzdá stimulu, niekedy aj bez zábran lietať.
Ak je štvornásobný reflex narušený, človek nemôže rýchlo prejsť z jedného druhu pohybu na druhý. V dôsledku toho sa štvorice podieľajú na organizácii dobrovoľných hnutí.
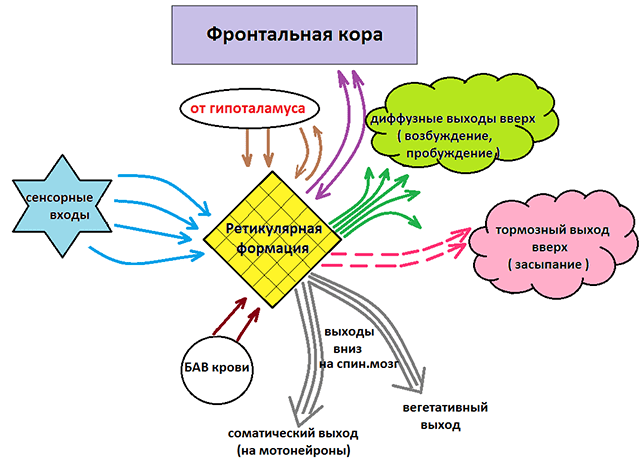
Retikulárna formácia mozgového kmeňa
Retikulárna formácia (formatioreticularis; RF) mozgu je reprezentovaná sieťou neurónov s početnými difúznymi spojeniami medzi sebou a s takmer všetkými štruktúrami centrálneho nervového systému. RF sa nachádza v hrúbke sivej hmoty predĺženej miechy, stredného mozgu a diencefala a je spočiatku spojená s RF miechy. V tomto smere je vhodné považovať to za jednotný systém... Sieťové spojenia neurónov RF medzi sebou umožnili Deitersovi nazvať to retikulárna formácia mozgu.
RF má priame a spätné spojenie s mozgovou kôrou, bazálnymi gangliami, diencefalom, mozočkom, stredom, predĺženou miechou a miechou.
Hlavnou funkciou RF je regulácia úrovne aktivity mozgovej kôry, mozočka, talamu a miechy.
Na jednej strane zovšeobecnená povaha vplyvu RF na mnohé mozgové štruktúry dávala dôvod považovať ho za nešpecifický systém. Štúdie so stimuláciou RF trupu však ukázali, že môže mať selektívne aktivačný alebo inhibičný účinok na rôzne formy správania, na senzorické, motorické a viscerálne systémy mozgu. Štruktúra siete zaisťuje vysokú spoľahlivosť RF fungovania, odolnosť voči škodlivým vplyvom, keďže lokálne poškodenie je vždy kompenzované zvyšnými sieťovými prvkami. Na druhej strane vysoká spoľahlivosť fungovania RF je zabezpečená tým, že podráždenie ktorejkoľvek jeho časti sa prejaví v činnosti celého RF danej konštrukcie difúznosťou spojov.
Väčšina RF neurónov má dlhé dendrity a krátky axón. Existujú obrovské neuróny s dlhým axónom, ktoré tvoria cesty z RF do iných oblastí mozgu, napríklad downstream, retikulospinálne a rubrospinálne. Axóny RF neurónov tvoria veľké množstvo kolaterál a synapsií, ktoré končia neurónmi v rôznych častiach mozgu. Axóny RF neurónov smerujúce do mozgovej kôry tu končia na dendritoch I a II vrstiev.
Aktivita RF neurónov je odlišná a v princípe je podobná aktivite neurónov v iných mozgových štruktúrach, no medzi RF neurónmi sú také, ktoré majú stabilnú rytmickú aktivitu nezávislú od prichádzajúcich signálov.
Zároveň sa v RF stredného mozgu a mostíka nachádzajú neuróny, ktoré sú v pokoji „tiché“, to znamená, že negenerujú impulzy, ale sú excitované pri stimulácii zrakových alebo sluchových receptorov. Ide o takzvané špecifické neuróny, ktoré poskytujú rýchlu reakciu na náhle, nerozpoznané signály. Značný počet RF neurónov je polysenzorických.
V RF medulla oblongata, stredného mozgu a mosta sa zbiehajú signály rôznych vnemov. Neuróny mostíka prijímajú signály hlavne zo somatosenzorických systémov. Signály z vizuálnych a sluchových senzorických systémov prichádzajú hlavne do RF neurónov stredného mozgu.
RF riadi prenos senzorickej informácie prechádzajúcej cez jadrá talamu, pretože pri intenzívnej vonkajšej stimulácii sú inhibované neuróny nešpecifických jadier talamu, čím sa odstraňuje ich inhibičný účinok z prenosových jadier toho istého talamu. a uľahčenie prenosu senzorických informácií do mozgovej kôry.
V RF pons, medulla oblongata, sú neuróny, ktoré reagujú na bolestivé podnety zo svalov alebo vnútorných orgánov, čo vytvára celkový difúzny diskomfort, nie vždy jasne lokalizovaný, bolestivý pocit „tupej bolesti“.
Opakovanie akéhokoľvek typu stimulácie vedie k zníženiu impulznej aktivity RF neurónov, t.j. adaptačné (závislosti) procesy sú vlastné RF neurónom mozgového kmeňa.
RF mozgového kmeňa priamo súvisí s reguláciou svalového tonusu, pretože RF mozgového kmeňa prijíma signály z vizuálnych a vestibulárnych analyzátorov a mozočku. Z RF do motorických neurónov miechy a jadier hlavových nervov sa prijímajú signály, ktoré organizujú polohu hlavy, trupu atď.
Retikulárne dráhy, ktoré uľahčujú činnosť motorických systémov miechy, pochádzajú zo všetkých častí RF. Dráhy vedúce z mostíka inhibujú aktivitu motorických neurónov miechy, ktoré inervujú flexorové svaly a aktivujú motorické neuróny extenzorových svalov. Dráhy vedúce z RF medulla oblongata majú opačné účinky. Podráždenie RF vedie k triaške, zvýšenému svalovému tonusu. Po ukončení dráždenia pretrváva ním spôsobený efekt ešte dlho, zrejme v dôsledku cirkulácie vzruchu v sieti neurónov.
Mozgový kmeň RF sa podieľa na prenose informácií z mozgovej kôry, miechy do mozočka a naopak z mozočka do rovnakých systémov. Funkciou týchto spojení je príprava a realizácia pohybových schopností spojených s habituáciou, orientačnými reakciami, reakciami na bolesť, organizáciou chôdze, pohybmi očí.
Regulácia autonómnej aktivity RF organizmu je popísaná v časti 4.3, tu však podotýkame, že táto regulácia sa najzreteľnejšie prejavuje vo fungovaní respiračných a kardiovaskulárnych centier. Pri regulácii autonómnych funkcií majú veľký význam tzv. štartovacie neuróny RF. Spôsobujú cirkuláciu excitácie v rámci skupiny neurónov, čím poskytujú tón regulovaných autonómnych systémov.
RF vplyvy možno široko rozdeliť na zostupné a vzostupné. Každý z týchto vplyvov má zasa inhibičný a stimulačný účinok.
Vzostupné účinky RF na mozgovú kôru zvyšujú jej tonus, regulujú excitabilitu jej neurónov, bez zmeny špecifickosti odpovedí na adekvátne stimuly. RF ovplyvňuje funkčný stav všetkých senzorických oblastí mozgu, preto hrá úlohu pri integrácii senzorických informácií z rôznych analyzátorov.
RF priamo súvisí s reguláciou cyklu bdelosti a spánku. Stimulácia niektorých RF štruktúr vedie k rozvoju spánku, zatiaľ čo stimulácia iných spôsobuje prebudenie. G. Magun a D. Moruzzi predložili koncept, podľa ktorého sa všetky typy signálov z periférnych receptorov dostávajú do medulla oblongata a pons cez RF kolaterály, kde sa prepínajú na neuróny, ktoré poskytujú vzostupné dráhy do talamu a potom do mozgová kôra.
Excitácia RF medulla oblongata alebo pons spôsobuje synchronizáciu aktivity mozgovej kôry, objavenie sa pomalých rytmov v jej elektrických parametroch a inhibíciu spánku.
RF excitácia stredného mozgu spôsobuje opačný efekt prebudenia: desynchronizáciu elektrickej aktivity kôry, objavenie sa rýchlych β-podobných rytmov s nízkou amplitúdou na elektroencefalograme.
G. Bremer (1935) ukázal, že ak je mozog prerezaný medzi predným a zadným tuberkulom štvorky, zviera prestane reagovať na všetky druhy signálov; ak sa rez uskutoční medzi predĺženou miechou a stredným mozgom (zatiaľ čo RF udržiava spojenie s predným mozgom), zviera reaguje na svetlo, zvuk a iné signály. V dôsledku toho je možné udržiavať aktívny analyzujúci stav mozgu pri zachovaní spojenia s predným mozgom.
Reakcia aktivácie mozgovej kôry sa pozoruje počas stimulácie RF medulla oblongata, stredného mozgu a diencefala. Dráždenie niektorých jadier talamu zároveň vedie ku vzniku ohraničených lokálnych oblastí vzruchu, a nie k jeho celkovému vybudeniu, ako je to pri podráždení iných častí RF.
RF mozgového kmeňa môže mať nielen vzrušujúci, ale aj inhibičný účinok na činnosť mozgovej kôry.
Zostupné vplyvy RF mozgového kmeňa na regulačnú aktivitu miechy stanovil I. M. Sechenov (1862). Ukázal, že keď je stredný mozog stimulovaný kryštálmi soli v žabe, reflexy stiahnutia labky sa objavujú pomaly, vyžadujú si intenzívnejšiu stimuláciu alebo sa neobjavia vôbec, to znamená, že sú inhibované.
G. Magun (1945-1950), ktorý spôsobuje lokálne podráždenie RF predĺženej miechy, zistil, že keď sú niektoré body podráždené, reflexy ohýbania prednej labky, kolena a rohovkového reflexu sú inhibované a spomalené. Keď bola RF stimulovaná v iných bodoch predĺženej miechy, rovnaké reflexy sa vyvolali ľahšie, boli silnejšie, t.j. uľahčila sa ich realizácia. Podľa Meguna iba RF medulla oblongata môže mať inhibičné účinky na reflexy miechy a facilitačné vplyvy sú regulované celým RF trupu a miechy.
9) (Cerebellum, jeho účasť na regulácii motorických a autonómnych funkcií)
Mozoček (cerebellum, malý mozog) je jednou z integračných štruktúr mozgu, ktorá sa podieľa na koordinácii a regulácii vôľových, mimovoľných pohybov, na regulácii autonómnych a behaviorálnych funkcií.
Vlastnosti morfofunkčnej organizácie a komunikácie cerebellum. Implementáciu týchto funkcií zabezpečujú nasledujúce morfologické znaky cerebellum:
1) mozočková kôra je stavaná celkom jednotne, má stereotypné spojenia, čo vytvára podmienky na rýchle spracovanie informácií;
2) hlavný nervový prvok kôry – Purkyňova bunka, má veľké množstvo vstupov a tvorí jediný axonálny výstup z mozočka, ktorého kolaterály končia na jeho jadrových štruktúrach;
3) do Purkyňových buniek sa premietajú takmer všetky typy zmyslových podnetov: proprioceptívne, kožné, zrakové, sluchové, vestibulárne atď.;
4) výstupy z mozočka zabezpečujú jeho spojenie s mozgovou kôrou, s kmeňovými útvarmi a miechou.
Mozoček je anatomicky a funkčne rozdelený na starú, starodávnu a novú časť.
Stará časť mozočka (archicerebellum) - vestibulárny mozoček - zahŕňa hrudkovito-flokulárny lalok. Táto časť má najvýraznejšie spojenie s vestibulárnym analyzátorom, čo vysvetľuje význam malého mozgu pri regulácii rovnováhy.
Staroveká časť mozočka (paleocerebellum) - spinálny mozoček - pozostáva z úsekov vermis a pyramídy mozočka, uvuly, pericelulárnej oblasti a prijíma informácie najmä z proprioceptívnych systémov svalov, šliach, periostu, kĺbových membrán.
Nový cerebellum (neocerebellum) zahŕňa kôru cerebelárnych hemisfér a oblasti červa; dostáva informácie z kôry, najmä pozdĺž frontocerebelárnej dráhy, z vizuálneho a sluchového receptívneho systému, čo naznačuje jeho účasť na analýze vizuálnych a sluchových signálov a organizácii reakcií na ne.
Mozočková kôra má špecifickú štruktúru, ktorá sa nikde v centrálnom nervovom systéme neopakuje. Horná (I) vrstva cerebelárnej kôry - molekulárna vrstva, pozostáva z paralelných vlákien, dendritických rozvetvení a axónov vrstiev II a III. V spodnej časti molekulárnej vrstvy sa nachádzajú košíky a hviezdicové bunky, ktoré zabezpečujú interakciu Purkyňových buniek.
Stredná (II) vrstva kôry je tvorená Purkyňovými bunkami, ktoré sú usporiadané v jednom rade a majú najvýkonnejší dendritický systém v centrálnom nervovom systéme. Na dendritickom poli jednej Purkyňovej bunky môže byť až 60 000 synapsií. V dôsledku toho tieto bunky vykonávajú úlohu zhromažďovania, spracovania a prenosu informácií. Axóny Purkyňových buniek sú jediným spôsobom, ktorým mozočkový kortex prenáša informácie do svojich jadier a jadra cerebelárnej štruktúry.
Pod II vrstvou kôry (pod Purkyňovými bunkami) sa nachádza zrnitá (III) vrstva pozostávajúca z buniek zŕn, ktorých počet dosahuje 10 miliárd.Axóny týchto buniek stúpajú nahor, delia sa v tvare T na povrch kôry, tvoriace dráhy kontaktov s Purkyňovými bunkami. Tu sú Golgiho bunky.
Z mozočku odchádzajú informácie cez hornú a dolnú časť nôh. Cez hornú časť nôh idú signály do talamu, mostíka, červeného jadra, jadier mozgového kmeňa a retikulárnej formácie stredného mozgu. Cez dolné končatiny mozočku signály idú do predĺženej miechy do jej vestibulárnych jadier, olív a retikulárnej formácie. Stredné pedikly cerebellum spájajú nový cerebellum s predným lalokom mozgu.
Impulzná aktivita neurónov sa zaznamenáva vo vrstve Purkyňových buniek a granulárnej vrstve a frekvencia generovania impulzov týchto buniek sa pohybuje od 20 do 200 za sekundu. Bunky cerebelárnych jadier generujú impulzy oveľa menej často - 1-3 impulzy za sekundu.
Stimulácia hornej vrstvy cerebelárneho kortexu vedie k dlhodobej (až 200 ms) inhibícii aktivity Purkyňových buniek. Rovnaká inhibícia sa vyskytuje pri svetelných a zvukových signáloch. Celkové zmeny v elektrickej aktivite mozočkovej kôry až po podráždenie zmyslového nervu akéhokoľvek svalového vzhľadu vo forme pozitívnej oscilácie (inhibícia aktivity kôry, hyperpolarizácia Purkyňových buniek), ktorá nastáva po 15-20 ms a trvá 20-30 ms, potom nastáva excitačná vlna v trvaní až 500 ms (depolarizácia Purkyňových buniek).
Signály z kožných receptorov, svalov, kĺbových membrán, periostu vstupujú do cerebelárnej kôry cez takzvané spinálno-cerebelárne dráhy: pozdĺž zadného (dorzálneho) a predného (ventrálneho). Tieto cesty k mozočku prechádzajú cez dolnú olivu medulla oblongata. Z olivových buniek pochádzajú takzvané popínavé vlákna, ktoré sa rozvetvujú na dendritoch Purkyňových buniek.
Jadrá mostíka posielajú aferentné cesty do mozočku, čím vytvárajú machové vlákna, ktoré končia v zrnových bunkách vrstvy III mozočkovej kôry. Medzi mozočkom a modrastým miestom medzimozgu je aferentné spojenie pomocou adrenergných vlákien. Tieto vlákna sú schopné difúzne uvoľňovať norepinefrín do medzibunkového priestoru cerebelárnej kôry, čím humorne menia stav dráždivosti jej buniek.
Axóny buniek vo vrstve III cerebelárneho kortexu spôsobujú inhibíciu Purkyňových buniek a buniek zŕn ich vlastnej vrstvy.
Purkyňove bunky zase inhibujú aktivitu neurónov v cerebelárnych jadrách. Mozočkové jadrá majú vysokú tonickú aktivitu a regulujú tonus množstva motorických centier v strednej, strednej, podlhovastej mieche.
Subkortikálny systém cerebellum pozostáva z troch funkčne odlišných jadrových útvarov: jadro stanu, korkové, guľovité a zubaté jadrá.
Stanové jadro prijíma informácie z mediálnej zóny cerebelárneho kortexu a je spojené s Deitersovým jadrom a RF medulla oblongata a stredného mozgu. Odtiaľ sa signály šíria retikulospinálnou dráhou k motorickým neurónom miechy.
Stredná cerebelárna kôra sa premieta na korkové a sférické jadrá. Z nich idú spojenia do stredného mozgu do červeného jadra, potom do miechy pozdĺž rubrospinálnej dráhy. Druhá cesta zo stredného jadra ide do talamu a ďalej do motorickej oblasti mozgovej kôry.
Zubaté jadro, ktoré dostáva informácie z laterálnej zóny cerebelárnej kôry, je spojené s talamom a cez neho - s motorickou zónou mozočkovej kôry.
Cerebelárna kontrola motorickej aktivity. Eferentné signály z mozočka do miechy regulujú silu svalových kontrakcií, poskytujú schopnosť predĺženej tonickej svalovej kontrakcie, schopnosť udržiavať optimálny svalový tonus v pokoji alebo počas pohybu, merať vôľové pohyby za účelom tohto pohybu, rýchlo prejsť z flexie do extenzie a naopak.
Cerebellum poskytuje synergiu kontrakcií rôznych svalov počas zložitých pohybov. Napríklad, keď človek urobí krok pri chôdzi, posunie nohu dopredu, zatiaľ čo ťažisko tela sa posunie dopredu za účasti chrbtových svalov. V prípadoch, keď cerebellum neplní svoju regulačnú funkciu, človek zažíva poruchy motorickej funkcie, čo je vyjadrené nasledujúcimi príznakmi.
1) asténia (asténia - slabosť) - zníženie sily svalovej kontrakcie, rýchla svalová únava;
2) astasia (astasia, z gréckeho a - nie, stasia - státie) - strata schopnosti predĺženej svalovej kontrakcie, ktorá sťažuje státie, sedenie atď.;
3) dystónia (distónia - porušenie tónu) - nedobrovoľné zvýšenie alebo zníženie svalového tonusu;
4) tremor (tremor - chvenie) - chvenie prstov, rúk, hlavy v pokoji; tento chvenie sa zvyšuje s pohybom;
5) dysmetria (dismetria - porušenie opatrenia) - porucha rovnomernosti pohybov, vyjadrená buď nadmerným alebo nedostatočným pohybom. Pacient sa pokúša vziať predmet zo stola a pritiahne ruku za predmet (hypermetria) alebo ju neprinesie k predmetu (hypometria);
6) ataxia (ataksia, z gréčtiny a - negácia, taksia - poriadok) - narušená koordinácia pohybov. Tu sa najzreteľnejšie prejavuje nemožnosť vykonávať pohyby v správnom poradí, v určitom poradí. Prejavmi ataxie sú aj adiadochokinéza, asynergia, opilecká kolísavá chôdza. Pri adiadochokinéze človek nie je schopný rýchlo otáčať dlaňami hore a dole. Pri asynergii svalov sa nedokáže bez pomoci rúk posadiť z polohy na bruchu. Opitá chôdza je charakteristická tým, že osoba kráča s nohami široko od seba a potáca sa zo strany na stranu od línie chôdze. Vrodených motorických úkonov u človeka nie je až tak veľa (napríklad sanie), ale väčšinu pohybov sa naučí počas života a stanú sa automatickými.
Fylogeneticky veľmi stará nervová štruktúra a dobre vyvinutá časť mozgového kmeňa plazov. Najprv to bola pomalá vodivá polysynaptická dráha úzko spojená s čuchovými a limbickými oblasťami. Postupná dominancia zraku a sluchu nad čuchom viedla k posunu zmyslových a motorických funkcií do tekta stredného mozgu. Priame dorzálne a teginálno-spinálne dráhy obchádzajú retikulárnu formáciu, ktorá je zodpovedná hlavne za autonómnu reguláciu. U cicavcov začalo tektum hrať sekundárnu úlohu pri prenose vzruchu po veľmi rýchlo vodivých vláknach spájajúcich mozgovú kôru s periférnymi motorickými a senzorickými neurónmi.
V ľudskom mozgu si retikulárna formácia zachováva spojenie s limbickým systémom a naďalej zohráva dôležitú úlohu v autonómnej a reflexnej regulácii.
Termín retikulárna formácia odkazujú len na polysynaptickú neurónovú sieť mozgového kmeňa, napriek tomu, že sieť siaha vpredu k talamu a hypotalamu a vzadu k propriospinálnemu traktu miechy.
Všeobecná štruktúra znázornené na obrázku nižšie. Stredný retikulárny útvar je tvorený množstvom šijacích jadier (grécky - nuclei raphe). Väčšina serotonergných dráh axiálnej časti nervového systému začína od jadier stehu.
Retikulárna formácia (RF).(A) Oddelenia. (B) Skupiny aminergných a cholinergných buniek.
Neďaleko sa nachádza paramediánna retikulárna formácia. Táto časť pozostáva výlučne z veľkých bunkových neurónov; v dolnej časti mosta a hornej časti medulla oblongata (až do úrovne fúzie retikulárnej formácie s centrálnym retikulárnym jadrom medulla oblongata) možno nájsť aj neuróny obrovských buniek.
Zvažuje sa najprednejšia časť laterálna malobunková retikulárna formácia... Dlhé dendrity malých bunkových neurónov sa v pravidelných intervaloch rozvetvujú. Dendrity majú prevažne priečny smer a cez medzery medzi nimi vedú dlhé cesty do talamu. Laterálny úsek tvoria prevažne aferentné neuróny. Sú pre nich vhodné vlákna zo všetkých citlivých dráh vrátane zmyslov.
Vlákna čuchu prechádzajú cez mediálny zväzok predného mozgu umiestnený vedľa hypotalamu.
Zrakové dráhy prechádzajú cez horný pahorok.
Sluchové vlákna vychádzajú z horného jadra olivy.
Vestibulárne vlákna pristupujú z mediálneho vestibulárneho jadra.
Somatické senzorické vlákna prechádzajú cez miechové retikulárne dráhy z miechy a vlastných (hlavného alebo hlavného mostíka) jadier trojklaného nervu.
Väčšina axónov neurónov malých buniek sa intenzívne rozvetvuje medzi dendritmi neurónov paramediánnej retikulárnej formácie. Niektoré z nich však tvoria synapsie s jadrami hlavových nervov a podieľajú sa na tvorbe pohybových programov.
Paramediálna retikulárna formácia- prevažne odvodný systém. Axóny sú relatívne dlhé, niektoré stúpajú nahor a tvoria synapsie s retikulárnou formáciou mozgového kmeňa alebo talamu. Z ostatných sa vetvia vzostupné aj zostupné vetvy, ktoré tvoria polysynaptickú sieť. Pre veľké bunkové neuróny sú vhodné vlákna z premotorickej kôry, z ktorých vznikajú retikulospinálne dráhy pons a medulla oblongata.

a) Aminergné neuróny mozgového kmeňa... Skupiny aminergných (alebo monoaminergných) neurónov rozptýlených pozdĺž retikulárnej formácie sú neuróny, ktorých mediátory sú tvorené z aromatických aminokyselín a majú množstvo účinkov na bunku. Jedna skupina produkuje neurotransmiter serotononín, tri ďalšie produkujú katecholamíny (dopamín, norepinefrín a adrenalín) a jedna skupina produkuje histamín.
 Serotonergné dráhy zo stredného mozgového kmeňa (šev).
Serotonergné dráhy zo stredného mozgového kmeňa (šev).
Serotonergné neuróny- najčastejšie neuróny v ktorejkoľvek časti centrálneho nervového systému (CNS). Patria sem neuróny stredného mozgu, ktorých vlákna stúpajú do mozgových hemisfér; pontínne neuróny rozvetvené do mozgového kmeňa a mozočku; bunky medulla oblongata, zostupujúce do miechy.
Všetky časti šedej hmoty centrálneho nervového systému sú preniknuté axonálnymi vetvami vylučujúcimi serotonín. Zvýšenie serotonergnej aktivity sa v klinickej praxi používa na liečbu takého bežného ochorenia, akým je veľká depresívna porucha.

Dopaminergné neuróny stredného mozgu sú zastúpené dvoma skupinami. Na mieste spojenia pneumatiky s nohami je čierna látka. Mediálne k nemu sú ventrálne jadrá tegmentálu, z ktorých sa rozširujú mezokortikálne vlákna do predného laloku a mezolimbické vlákna, ktoré idú priamo do nucleus accumbens.
Noradrenergné (norepinefrinergné) neuróny o niečo menej hojný ako serotonergný. Asi 90 % tiel neurónov je sústredených v locus ceruleus v spodnej časti IV komory na hornom konci mostíka. Cesty začínajú od modrého bodu vo všetkých smeroch, ako je znázornené na obrázku nižšie.
 Noradrenergné dráhy z mostíka a predĺženej miechy.
Noradrenergné dráhy z mostíka a predĺženej miechy.
Neuróny vylučujúce adrenalín (adrenalín vylučujúce). relatívne málo a nachádzajú sa hlavne v rostrálnych/kaudálnych oblastiach medulla oblongata. Jedna časť vlákien stúpa do hypotalamu, druhá smeruje nadol a vytvára synapsie s pregangliovými sympatickými neurónmi miechy.
V mozgových hemisférach je iónová a elektrická aktivita aminergných neurónov výrazne odlišná. Po prvé, pre každý amín existuje viac ako jeden typ postsynaptických receptorov. Po druhé, uvoľňujú sa aj niektoré aminergné neuróny bielkovinové látky, schopný regulovať činnosť mediátora – spravidla zvyšuje jeho trvanie. Po tretie, väčšie kortikálne neuróny dostávajú veľa excitačných a inhibičných vplyvov z lokálnych cirkulujúcich excitačných sietí a majú tiež mnoho rôznych typov receptorov. Aktivácia jedného typu aminergného receptora môže viesť k silnému alebo slabému účinku v závislosti od počiatočného excitovaného stavu neurónu.
Naše znalosti o fyziológii a farmakodynamike aminergných neurónov nie sú ani zďaleka úplné, ale ich význam v širokej škále funkcií správania je nepochybný.
Časť prierezu hornou časťou mosta zobrazujúca prvky retikulárnej formácie.
Zložitá štruktúra ľudského mozgu odhaľuje tajomstvá nášho správania, vysvetľuje zákonitosti duševnej činnosti, prúdenie emócií a pocitov. Každá hemisféra mozgu je zodpovedná za svoje špecifické funkcie a úlohy (napríklad je známe, že pravá je zodpovedná za logiku a ľavá za predstavivosť a fantáziu), existujú však aj štruktúry, ktoré poskytujú jednotný a koordinovaný prácu celého centrálneho nervového systému. Jednou z týchto štruktúr je retikulárna formácia.
Všeobecné informácie
Retikulárna formácia je časť mozgového kmeňa, ktorú predstavuje rozsiahla sieť nervových buniek a jadier, ktoré spájajú rôzne časti mozgu. Na rozdiel od iných štruktúr - napríklad talamus, hypotalamus, cerebellum - ktoré majú určitý celistvý tvar (jadrá, žľazy), retikulárna formácia nie je reprezentovaná jedným morfologickým útvarom, ale je "sieťou" (z latinského retikulum - sieť) dendritov a axónov, ktoré s rôznym stupňom hustoty prenikajú medzi oddelenia a štruktúry mozgu, spájajú ich navzájom a zabezpečujú ich spoločnú aktivitu.
Metaforicky povedané: ak je náš mozog prezentovaný vo forme nejakého produktu, povedzme košele, potom retikulárny útvar sú vlákna, ktorými je košeľa šitá. Retikulárna formácia preniká do štruktúr predĺženej miechy, stredného mozgu a mosta, má priame spojenie s mozočkom, miechou, talamom a nepriamo aj s nadložnými úsekmi: hypotalamom, zrakovými jadrami a kôrou.
Ako to funguje
Retikulárna formácia zahŕňa obrovské množstvo neurónov s rozvetvenými dendritmi a dlhými axónmi, vďaka ktorým je možné prenášať nervové impulzy do rôznych častí mozgu a miechy. V tomto prípade možno rozlíšiť dve najväčšie skupiny nervových zhlukov:
- Retikulotegmentálne jadro, ktorého neuróny prijímajú signály z nadložných úsekov GM (štvornásobok, talamus) a prenášajú ich ďalej do cerebelárnych štruktúr, čím regulujú niektoré životne dôležité motorické funkcie: koordináciu pohľadu, pohyby očí.
- Na regulácii dýchania a cievnej inervácie sa podieľa laterálne jadro, ktorého neuróny vystupujú zo štruktúr miechy a vestibulárnych jadier a poskytujú GM kortexu informácie o polohe tela v priestore.
- Okrem toho retikulárna formácia zahŕňa neuróny, ktoré sa významnou mierou podieľajú na práci centier termoregulácie, sýtosti a hladu.

Hlavné funkcie
Hlavným účelom retikulárnej formácie je senzorická analýza mnohých signálov z rôznych častí GM.
Vďaka úzkemu spojeniu s miechou sa aktívne podieľa aj na motorickej regulácii, od prehĺtacieho reflexu až po zložité motorické operácie. Okrem toho retikulárna formácia má aktivačný účinok na celý GM ako celok, pričom sa podieľa na regulácii cyklov spánku a bdenia.
Vo všeobecnosti sú funkcie retikulárnej formácie nasledovné:
- Regulácia kostrových svalov (zapojených do riadenia pohybov tela) a autonómnych funkcií (dýchanie, kýchanie, krvný obeh atď.).
- Kontrola procesov spánku a bdenia (kvôli poskytovaniu aktivačného a inhibičného účinku na mozgovú kôru).
- Aktivačná funkcia (prejavuje sa tým, že retikulárna formácia poskytuje neustálu tonickú stimuláciu GM kôry, vďaka čomu je možné udržať pozornosť, vedomie a priebeh myšlienkových procesov.)
- Spracovanie signálov z vonkajšieho a vnútorného prostredia.
Charakteristické črty práce retikulárnej formácie sú spojené predovšetkým s jej určitými vlastnosťami:
Nervové bunky, ktoré tvoria retikulárnu formáciu, majú zvýšenú kapacitu tonickej excitácie. To znamená, že väčšina neurónov je v neustálom vzrušení a vytvára nervové impulzy, ktoré sa prenášajú do nadložných častí GM. Táto tonická aktivita je spôsobená niekoľkými faktormi:
- Priepustnosť veľkého počtu signálov cez štruktúry retikulárnej formácie. Tu je jednoduchá analógia: predstavte si klavír alebo nejaký iný strunový nástroj. Je jasné, že keď sa struny priamo dotkneme, začnú vibrovať a vydávať zvuk. To isté sa deje s nervovými bunkami, keď k nim prichádzajú signály z iných neurónov. Predstavme si však ďalej, že sa nedotýkame priamo strún nástroja, ale povedzme skáčeme vedľa neho a silno búchame nohami o podlahu. . Zvuk nástroja síce nepočujeme, no aj tak bude prebiehať sotva badateľné chvenie strún. To isté sa deje s neurónmi retikulárnej formácie. Pretože cez ňu neustále prechádzajú niektoré signály (aferentné aj eferentné) z rôznych štruktúr centrálneho nervového systému, vytvára to konštantnú tonickú excitáciu neurónov retikulárnej formácie, pretože je v epicentre konštantnej neuronovej sústavy. - výmena impulzov.
- Zvýšená citlivosť neurónov na chemikálie (hormóny, lieky, psychotropné látky). Ráno vypitá šálka kávy „zapína“ štruktúry retikulárnej formácie a vďaka dlhodobému zachovaniu vzruchu v jej neurónoch nás udržiava aktívnych.
Vplyv RF smerom nadol a nahor
Ako už bolo uvedené, retikulárna formácia má vzrušujúci a inhibičný účinok na rôzne časti GM. V tomto prípade možno rozlíšiť dve oddelenia, ktoré sa špecializujú na prenos vzruchu do určitých štruktúr mozgu.
Zostupné oddelenie: reprezentované autonómnymi a motorickými centrami a má zostupný vplyv na oddelenia miechy. Zodpovedajúce nervové zhluky regulujú činnosť respiračných, vazomotorických, slinných centier, ako aj centier zodpovedných za výstavbu jednoduchých a zložitých motorických reakcií. To naznačuje rozhodujúcu úlohu centrálneho nervového systému pri regulácii aj elementárnych nepodmienených reflexov. Stimulácia zostupného úseku vedie k inhibícii centier chrbtice a navodzuje v prirodzenom prostredí stav hlbokého spánku (spánok „bez zadných nôh“). Rovnaký efekt možno navodiť umelo, napríklad uvedením človeka do stavu tranzu alebo anestézie.
Vzostupný úsek: reprezentovaný nervovými vláknami spájajúcimi štruktúry retikulárnej formácie s nadložnými úsekmi: talamus, hypotalamus, cerebellum a kôra. Vplyv nahor pôsobí stimulačne na kortikálne štruktúry a poskytuje aktívny stav vedomia. Vplyv nahor sa nezastaví ani keď spíme. Ak by sa náš mozog mohol úplne „vypnúť“, každé prebudenie by sa podobalo pôrodu: kto som? Kde som? Ako som sa sem dostal? Vďaka práci retikulárnych štruktúr však stále máme možnosť vrátiť sa vždy do toho počiatočného stavu vedomia, v ktorom sme boli pred chvíľou spánku. Navyše, aj počas nočného odpočinku máme stále možnosť reagovať na niektoré životne dôležité podnety, t.j. my spravidla nespíme „mŕtvy spánok“ a môžeme sa zobudiť, ak sa dieťa pohne a plakalo v blízkosti, niečo hlasno spadlo atď.
Prejav poškodenia konštrukcií
Retikulárna formácia hrá významnú úlohu v integračnej aktivite celého mozgu. Vzhľadom na výkon funkcie vedúceho vodiča všetkých druhov nervových vzruchov do všetkých častí centrálneho nervového systému je retikulárna formácia v neustálej prevádzke. Nadmerné duševné a emocionálne preťaženie je škodlivé pre mozog vo všeobecnosti a najmä pre retikulárnu formáciu. Našťastie včasné podanie sedatív môže (vzhľadom na zvýšenú náchylnosť neurónov na chemické účinky) rýchlo napraviť situáciu a normalizovať stav.
Možné sú však aj menej priaznivé výsledky. Poškodenie je možné v dôsledku kraniocerebrálnej traumy, onkologických ochorení mozgu, infekčných lézií.
Hlavným prejavom nešťastia je strata vedomia.
Porušenie vzostupných spojení sa prejavuje v stave apatie, slabosti, zvýšenej ospalosti, motorickej dezintegrácie, narušeného nočného spánku. Časté sú sprievodné autonómne poruchy.
Retikulárna formácia - súbor rôznych, umiestnených pozdĺž mozgového kmeňa, majúcich aktivačný alebo inhibičný účinok na rôzne štruktúry centrálneho nervového systému, čím riadia ich reflexnú aktivitu.
Retikulárna formácia mozgového kmeňa má aktivačný účinok na bunky a inhibičný účinok na motorické neuróny miechy. Vysielaním inhibičných a excitačných impulzov do miechy k jej motorickým neurónom sa retikulárna formácia podieľa na regulácii tonusu kostrového svalstva.
Retikulárna formácia udržuje tonus autonómnych centier, integruje sympatické a parasympatické vplyvy, prenáša modulačný účinok z hypotalamu a mozočka na vnútorné orgány.
Funkcie retikulárnej formácie
Somatomotorické ovládanie(aktivácia kostrového svalstva), môže byť priamo cez tr. reticulospinalis a nepriame cez, olivy, tuberkulózy štvorce, červené jadro, čierna hmota, striatum, talamické jadrá a dokonca somatomotorické zóny kôry.
Somatosenzitívna kontrola, t.j. pokles hladín somatosenzorických informácií - "pomalá bolesť", modifikácia vnímania rôznych druhov zmyslovej citlivosti (sluch, zrak, vestibulácia, čuch).
Visceromotorické ovládanie stav kardiovaskulárneho, dýchacieho systému, činnosť hladkých svalov rôznych vnútorných orgánov.
Neuroendokrinná transdukcia cez vplyv na neurotransmitery, centrá hypotalamu a potom hypofýzu.
Biorytmy prostredníctvom spojení s hypotalamom a epifýzou.
Rôzne funkčné stavy tela(spánok, prebudenie, stav vedomia, správanie) sa uskutočňujú prostredníctvom početných spojení jadier retikulárnej formácie so všetkými časťami centrálneho nervového systému.
Koordinácia práce rôznych centrá mozgového kmeňa poskytovanie komplexných viscerálnych reflexných reakcií (kýchanie, kašeľ, vracanie, zívanie, žuvanie, sanie, prehĺtanie atď.).
Štruktúra retikulárnej formácie
Retikulárna formácia tvorené súborom početných neurónov ležiace oddelene alebo zoskupené do jadier (pozri obr. 1 a 2). Jeho štruktúry sú lokalizované v centrálnych oblastiach trupu, začínajúc od horných segmentov krčnej miechy po hornú úroveň mozgového kmeňa, kde sa postupne spájajú s jadrovými skupinami. Retikulárna formácia zaberá priestory medzi jadrami hlavových nervov, inými jadrami a dráhami prechádzajúcimi mozgovým kmeňom.
Neuróny retikulárnej formácie sa vyznačujú širokou škálou tvarov a veľkostí, ale ich spoločným znakom je, že tvoria početné synaptické kontakty s dlhými dendritmi a široko sa vetviacimi axónmi, a to ako medzi sebou, tak aj s neurónmi v iných mozgových jadrách. Tieto pobočky tvoria akúsi sieť ( retikulum), odkiaľ pochádza názov - retikulárny útvar. Neuróny, ktoré tvoria jadrá retikulárnej formácie, majú dlhé axóny ktoré tvoria cesty do miechy, jadier mozgového kmeňa a iných oblastí mozgu.

Ryža. 1. Najdôležitejšie štruktúrne útvary stredného mozgu (prierez)
Neuróny retikulárnej formácie dostávajú početné aferentné signály z rôznych štruktúr centrálneho nervového systému. Existuje niekoľko skupín neurónov, ktoré prijímajú tieto signály. to skupina neurónov laterálneho jadra retikulárna formácia, lokalizovaná v medulla oblongata. Jadrové neuróny prijímajú aferentné signály z interneurónov miechy a sú súčasťou jednej z nepriamych spinocerebelárnych dráh. Okrem toho prijímajú signály z vestibulárnych jadier a dokážu integrovať informácie o stave aktivity interneurónov spojených s motorickými neurónmi miechy a o polohe tela a hlavy v priestore.
Ďalšia skupina je neuróny retikulotegmentálneho jadra nachádza sa na hranici dorzálneho okraja mosta. Dostávajú aferentné synaptické vstupy z neurónov pretektálnych jadier a horných kopčekov štvorice a posielajú svoje axóny do štruktúr mozočka, ktoré sa podieľajú na kontrole pohybov očí.
Neuróny retikulárnej formácie dostávajú rôzne signály prostredníctvom dráh, ktoré ich spájajú s mozgovou kôrou (kortikoretikulospinálne dráhy), substantia nigra atď.

Ryža. 2. Umiestnenie niektorých jadier v mozgovom kmeni a hypotalame: 1 - paraventrikulárne; 2 - dorzomediálne: 3 - preoptické; 4 - supraoptika; 5 - späť
Okrem opísaných aferentných dráh sú signály vysielané do retikulárnej formácie cez axonálne kolaterály dráhy zmyslových systémov. Zároveň sa signály z rôznych receptorov (hmatový, zrakový, sluchový, vestibulárny, bolestivý, teplotný, proprioreceptory, receptory vnútorných orgánov) môžu zbiehať k tomu istému.
Z vyššie uvedeného zoznamu hlavných aferentných spojení retikulárnej formácie s inými oblasťami centrálneho nervového systému možno vidieť, že stav jej tonickej neuronálnej aktivity je determinovaný prílevom takmer všetkých typov senzorických signálov zo senzorických neurónov, ako aj signály z väčšiny štruktúr centrálneho nervového systému.
Klasifikácia retikulárnej formácie v závislosti od smeru vlákien|
oddelenia |
Charakteristický |
|
Zostupné oddelenie |
Vegetatívne centrá:
Motorové centrá:
|
|
Vzostupné oddelenie |
Retikulotalamický Retikulohypotalamické Retikulocerebelárne Retikulokortikálna: aktivujúca; hypnogénny |
Jadrá retikulárnej formácie a ich funkcie
Dlho sa verilo, že retikulárna formácia, ktorej štruktúra je charakterizovaná širokými interneuronálnymi spojeniami, integruje signály rôznych modalít bez vysielania špecifických informácií. Čoraz viac sa však ukazuje, že retikulárny útvar je nielen morfologicky, ale aj funkčne heterogénny, hoci rozdiely medzi funkciami jeho jednotlivých častí nie sú také zjavné, ako je to typické pre iné oblasti mozgu.
V skutočnosti mnohé neurónové skupiny retikulárnej formácie tvoria jej jadrá (centrá), ktoré vykonávajú špecifické funkcie. Sú to neurónové skupiny, ktoré sa tvoria vazomotorické centrum medulla oblongata (obrovské bunkové, paramediálne, laterálne, ventrálne, kaudálne jadrá medulla oblongata), dýchacie centrum(obrovská bunka, malé bunkové jadro medulla oblongata, orálne a chvostové jadro mostíka), žuvacie centrá a prehĺtanie(laterálne, paramediálne jadro medulla oblongata), centrá pohybu očí(paramediálna časť mosta, rostrálna časť stredného mozgu), centrá regulácie svalového tonusu(rostrálne jadro ponsu a kaudálne jadro medulla oblongata) atď.
Jednou z najdôležitejších nešpecifických funkcií retikulárnej formácie je regulácia všeobecnej nervovej aktivity kôry a iné štruktúry centrálneho nervového systému. V retikulárnej formácii sa hodnotí biologický význam prichádzajúcich senzorických signálov a v závislosti od výsledkov tohto hodnotenia môže aktivovať alebo inhibovať prostredníctvom nešpecifických alebo špecifických neurónových skupín talamu nervové procesy v celej mozgovej kôre alebo vo všetkých jednotlivé zóny. Preto sa kmeňový retikulárny útvar nazýva aj tzv systém aktivácie suda mozog. Vďaka týmto vlastnostiam môže retikulárna formácia ovplyvniť úroveň celkovej aktivity kôry, ktorej udržanie je najdôležitejšou podmienkou pre udržanie vedomia, stavu bdelosti a formovania ohniska pozornosti.
Zvýšenie aktivity retikulárnej formácie (oproti všeobecnému vysokému pozadiu) v jednotlivých senzorických, asociačných oblastiach kôry poskytuje schopnosť izolovať a spracovať špecifické informácie, ktoré sú v danom čase pre telo najdôležitejšie a organizovať adekvátnu reakciu správania. reakcie. Zvyčajne týmto reakciám, organizovaným za účasti retikulárnej formácie mozgového kmeňa, predchádzajú orientačné pohyby očí, hlavy a tela v smere zdroja signálu, zmeny dýchania a krvného obehu.
Aktivačný účinok retikulárnej formácie na kôru a ďalšie štruktúry centrálneho nervového systému sa uskutočňuje pozdĺž vzostupných dráh vychádzajúcich z obrovských buniek, laterálnych a ventrálnych retikulárnych jadier medulla oblongata, ako aj z jadier ponsu. a stredný mozog. Po týchto dráhach sú prúdy nervových impulzov vedené do neurónov nešpecifických jadier talamu a po ich spracovaní sú v jadrách talamu prepínané na následný prenos do kôry. Okrem toho sú z uvedených retikulárnych jadier vedené toky signálov do neurónov zadného hypotalamu a bazálnych ganglií.
Okrem regulácie nervovej aktivity vyšších častí mozgu môže retikulárna formácia regulovať zmyslové funkcie. Deje sa tak ovplyvňovaním vedenia aferentných signálov do nervových centier, excitability neurónov nervových centier, ako aj citlivosti receptorov. Zvýšenie aktivity retikulárnej formácie je sprevádzané zvýšením aktivity neurónov sympatického nervového systému, ktorý inervuje zmyslové orgány. V dôsledku toho sa môže zvýšiť zraková ostrosť, sluch a hmatová citlivosť.
Spolu so vzostupnými aktivačnými a inhibičnými vplyvmi na vyššie časti mozgu sa podieľa aj retikulárna formácia. regulácia pohybov s aktivačnými a inhibičnými účinkami na miechu. Na jeho jadrách sa prepínajú ako vzostupné dráhy z proprioceptorov a miechy do mozgu, tak zostupné motorické dráhy z mozgovej kôry, bazálnych jadier, mozočka a červeného jadra. Vzostupné nervové dráhy vedúce z retikulárnej formácie do talamu a kôry síce zohrávajú úlohu najmä pri udržiavaní celkovej úrovne aktivity mozgových hemisfér mozgu, no práve táto ich funkcia je dôležitá pre plánovanie bdelej kôry , spúšťať, vykonávať pohyby a kontrolovať ich vykonávanie. Medzi vzostupnými a zostupnými dráhami je cez retikulárnu formáciu veľké množstvo kolaterálnych spojení, prostredníctvom ktorých sa môžu vzájomne ovplyvňovať. Existencia takejto úzkej interakcie vytvára podmienky pre vzájomné ovplyvňovanie oblasti retikulárnej formácie, ktorá prostredníctvom talamu ovplyvňuje činnosť kôry, plánovanie a spúšťanie pohybov, a oblasti retikulárnej formácie, ktorá ovplyvňuje výkonné nervové mechanizmy kôry. miecha. V retikulárnej formácii existujú skupiny neurónov, ktoré posielajú väčšinu axónov do cerebellum, ktorý sa podieľa na regulácii a koordinácii zložitých pohybov.
Pozdĺž zostupných retikulospinálnych dráh retikulárna formácia priamo ovplyvňuje funkcie miechy. Priamy vplyv na jeho motorické centrá sa vykonáva mediálny retikulospinálny trakt, prichádzajúce z jadier mosta a aktivujúce hlavne inter- a y-motorické neuróny extenzorov a inhibujúce motorické neuróny flexorových svalov trupu a končatín. Autor: laterálny retikulospinálny trakt vychádzajúc z obrovského bunkového jadra medulla oblongata, retikulárna formácia má aktivačný účinok na inter- a y-motorické neuróny flexorových svalov končatín a inhibuje neuróny extenzorových svalov.
Z experimentálnych pozorovaní na zvieratách je známe, že stimulácia viac rostrálne umiestnených neurónov retikulárnej formácie na úrovni medulla oblongata a stredného mozgu má difúzny facilitačný účinok na miechové reflexy a stimuláciu neurónov v kaudálnej časti medulla oblongata. je sprevádzaná inhibíciou synálnych reflexov.
Aktivačný a inhibičný účinok retikulárnej formácie na motorické centrá miechy sa môže uskutočniť prostredníctvom y-motorických neurónov. V tomto prípade retikulárne neuróny rostrálnej oblasti retikulárnej formácie aktivujú y-motorické neuróny, ktoré inervujú intrafuzálne svalové vlákna, spôsobujú ich kontrakciu, aktivujú receptory svalového vretienka. Tok signálov z týchto receptorov aktivuje a-motoneuróny a spôsobí kontrakciu príslušného svalu. Neuróny v kaudálnej oblasti retikulárnej formácie inhibujú aktivitu y-motorických neurónov miechy a spôsobujú svalovú relaxáciu. Rozloženie tonusu vo veľkých svalových skupinách závisí od rovnováhy nervovej aktivity v týchto oblastiach retikulárnej formácie. Keďže táto rovnováha závisí od zostupných vplyvov na retikulárnu formáciu mozgovej kôry, bazálnych ganglií, hypotalamu, mozočka, tieto mozgové štruktúry môžu tiež ovplyvniť distribúciu svalového tonusu a držanie tela cez retikulárnu formáciu a iné jadrá mozgového kmeňa.
Široké rozvetvenie axónov retikulospinálnych dráh v mieche vytvára podmienky pre vplyv retikulárnej formácie na takmer všetky motorické neuróny, a teda na stav svalov rôznych častí tela. Táto vlastnosť poskytuje efektívny efekt retikulárnej formácie na reflexné rozloženie svalového tonusu, držanie tela, orientáciu hlavy a tela v smere vonkajších podnetov a účasť retikulárnej formácie na realizácii vôľových svalových pohybov v proximálnych partiách. tela.
V centrálnej časti jadra retikulárnej obrovskej bunky je miesto, ktorého podráždenie inhibuje všetky motorické reflexy miechy. Prítomnosť takejto inhibície mozgových štruktúr na mieche objavil I.M. Sechenov v pokusoch na žabách. Podstatou experimentov bolo štúdium stavu miechových reflexov po pretínaní mozgového kmeňa na úrovni diencefala a podráždení kaudálnej časti rezu kryštálom chloridu sodného. Ukázalo sa, že motorické miechové reflexy sa počas stimulácie neobjavili alebo sa oslabili a obnovili po odstránení podráždenia. Najprv sa teda zistilo, že jedno nervové centrum môže inhibovať aktivitu druhého. Tento jav bol pomenovaný centrálne brzdenie.
Retikulárna formácia zohráva významnú úlohu v regulácii nielen somatických, ale aj autonómnych funkcií (retikulárne jadrá mozgového kmeňa sú súčasťou štruktúry životne dôležitých častí dýchacieho centra a centier regulácie krvného obehu). Vzniká laterálna skupina mostonosných retikulárnych jadier a dorzolaterálne tegmentálne jadro močové centrum mosta. Axóny neurónov jadier tohto centra dosahujú pregangliové neuróny sakrálnej miechy. Stimulácia neurónov týchto jadier v pons je sprevádzaná kontrakciou svalstva steny močového mechúra a močením.
V dorzolaterálnom mostíku sa nachádza parabrachiálne jadro, na ktorého neurónoch končia vlákna senzorických neurónov chuti. Neuróny jadra, podobne ako neuróny modrastej škvrny a substantia nigra, obsahujú neuromelanín. Počet takýchto neurónov v parabrachiálnom jadre klesá pri Parkinsonovej chorobe. Neuróny parabrachiálneho jadra majú spojenie s neurónmi hypotalamu, amygdaly, jadier stehov, osamelého traktu a iných jadier mozgového kmeňa. Predpokladá sa, že parabrachiálne jadrá súvisia s reguláciou autonómnych funkcií a pokles ich počtu pri parkinsonizme vysvetľuje výskyt autonómnych porúch pri tomto ochorení.
Pri pokusoch na zvieratách sa ukázalo, že pri podráždení niektorých lokálnych oblastí retikulárnych štruktúr medulla oblongata a pons môže dôjsť k inhibícii aktivity kôry a spánku. V tomto prípade sa na EEG objavia nízkofrekvenčné (1-4 Hz) vlny. Na základe popísaných skutočností sa predpokladá, že najdôležitejšími funkciami vzostupných vplyvov retikulárnej formácie sú regulácia cyklu spánok-bdenie a úroveň vedomia. Ukázalo sa, že množstvo jadier retikulárnej formácie mozgového kmeňa priamo súvisí so vznikom týchto stavov.
Takže na každej strane centrálnej sutúry mostíka sú paramediálne retikulárne jadrá, príp. jadro stehu obsahujúce serotonergné neuróny. V kaudálnej časti mosta k nim patrí dolné centrálne jadro, ktoré je pokračovaním jadra sutúry predĺženej miechy a v rostrálnej časti mostíka horné centrálne jadro, nazývané ankylozujúca spondylitída, resp. stredové jadro švu, je súčasťou jadier ponsového švu.
Skupina jadier sa nachádza v rostrálnej časti pons na dorzálnej strane opercula modrastá škvrna. Obsahujú asi 16 000 – 18 000 noradrenergných neurónov obsahujúcich melanín, ktorých axóny sú široko zastúpené v rôznych častiach centrálneho nervového systému – hypotalame, hipokampe, mozgovej kôre, mozočku a mieche. Modrastá škvrna zasahuje do stredného mozgu a jej neuróny možno vysledovať v priestore stredného mozgu. Počet neurónov v jadre modrastej škvrny klesá pri parkinsonizme, Alzheimerovej chorobe a Downovom syndróme.
Sérotonergné aj norepinefrínové neuróny v retikulárnej formácii zohrávajú úlohu pri riadení cyklu spánku a bdenia. Potlačenie syntézy serotonínu v jadrách švu vedie k rozvoju nespavosti. Predpokladá sa, že serotonergné neuróny sú súčasťou neurónovej siete, ktorá reguluje spánok s pomalými vlnami. Pôsobením serotonínu na neuróny modrastej škvrny dochádza k paradoxnému spánku. Zničenie jadier modrastej škvrny u pokusných zvierat nevedie k rozvoju nespavosti, ale spôsobuje vymiznutie paradoxnej fázy spánku na niekoľko týždňov.
Sieť nervových buniek umiestnených v strede mozgového kmeňa opísal maďarský anatóm Jozsef Lenhossek už v roku 1855. Fylogeneticky ide o starodávnu skupinu buniek, ktorú majú všetky stavovce a zohráva dôležitú úlohu pri regulácii činnosti centrálneho nervového systému. ,
V strede sú retikulárne bunky; po stranách - zmyslové dráhy. U ľudí začína retikulárna formácia nad miechou v predĺženej mieche a cez mostík a stredný mozog sa tiahne k diencephalonu. Zvláštnosťou tohto útvaru je, že je laterálne (laterálne) obklopený zmyslovými dráhami popísanými "v prvej časti knihy. Vlákna retikulárnej formácie teda ležia v strede a zvonku sú obklopené vrstvou zmyslových dráh. Neurónová sieť retikulárnej formácie je veľmi heterogénna: veľkosť neurónov sa pohybuje od 10 do 100 mikrónov a sú tiež umiestnené rôznymi spôsobmi. V retikulárnej formácii je možný rýchly aj pomalý prenos impulzov. väčšina autorov nemá žiadnu organizáciu: v tejto difúznej predĺženej formácii nemožno rozlíšiť žiadne konkrétne štruktúry. Nedávno však výskumníci navrhli prítomnosť samostatných skupín buniek v retikulárnej formácii. Všetci však stále súhlasia s tým, že rozdiely sú iba funkčné: v niektorých oblastiach sa nachádzajú inhibičné jadrá av iných - aktivačné jadrá.
Viacnásobné pripojenia. Kahal ako prvý dospel k záveru, že všetky zmyslové dráhy z miechy a predĺženej miechy do kôry vydávajú kolaterály do aktivujúceho retikulárneho systému. Pre aktiváciu retikulárnej siete sú tieto kolaterály mimoriadne dôležité. Kolaterály k retikulárnej formácii tiež idú z kôry, talamu a mozočku. Retikulárna formácia tiež posiela vlákna do kôry, ako aj do motorických neurónov miechy.
"Bdelé neuróny". Funkciu aktivačnej retikulárnej formácie prvýkrát preukázali experimenty Maguna a Moruzziho v roku 1949. Stimulovali retikulárny systém prostredníctvom elektród implantovaných do mozgového kmeňa, a tak vyvolali prebúdzaciu reakciu, teda prebudili spiace zviera. Zároveň bolo možné na obrázku EEG pozorovať prechod z alfa aktivity na beta. Správanie zvieraťa sa zodpovedajúcim spôsobom zmenilo.
Vzostupný aktivačný systém. Je zaujímavé, že prebudenie môže byť spustené izolovanou stimuláciou retikulárneho systému, aj keď sú senzorické dráhy prerušené. Medzitým v opačnom prípade - po poškodení retikulárnej formácie vysokofrekvenčným elektrickým prúdom, so všetkými zmyslovými dráhami neporušenými - žiadna stimulácia neprebudí zviera z hlbokého spánku. Zistilo sa, že objav Meguna a Moruzziho súvisí s Bremerovým pozorovaním opísaným vyššie a s Pavlovovým orientačným reflexom. Systém v mozgovom kmeni, podporujúci
tonus mozgovej kôry, Magun nazval vzostupný aktivačný systém (obr. 37).
Prebudenie, ale nie informácie. Stupeň retikulárnej aktivity je určený výlučne impulzmi vstupujúcimi do retikulárnej formácie pozdĺž kolaterál senzorických dráh, t.j. vonkajších stimulov. Retikulárna formácia je zbavená vlastnej spontánnej aktivity. Jeho funkciou je „prebudiť“ celú masu neurónov v oboch mozgových hemisférach. Udržiavať bdelosť je,
Somatosenzorické
kôra ^ Thalamus
pravdepodobne funkcia kôry; retikulárna formácia "zahŕňa" iba kortikálne neuróny. Ide o všeobecný „poplachový systém“, ktorý reaguje na všetky zmyslové impulzy rovnako, bez ohľadu na to, či pochádzajú zo sluchových, zrakových alebo kožných receptorov. Jeho funkciou je prebudiť kôru, nie sprostredkovať konkrétne informácie.
Spustite poplachovú reakciu. Je veľmi pravdepodobné, že aktivujúca retikulárna formácia je spojená aj s endokrinnými mechanizmami. Napríklad po zavedení adrenalínu experimentálnym zvieratám možno retikulárnu aktiváciu posúdiť pomocou desynchronizácie EEG. Ukázalo sa, že adrenalín, teda zvýšená aktivita sympatiku, vždy spôsobuje retikulárnu aktiváciu, ktorá zlepšuje stav bdelosti zvieraťa. Cannon nazval výslednú komplexnú zmenu telesných funkcií poplachovou reakciou. Tento stav sa prejavuje zrýchlením
srdcová frekvencia, zvýšený krvný tlak a telesná teplota, hyperglykémia a pod. Aktivačný retikulárny systém teda môže sám o sebe spôsobiť zvýšenie tonusu sympatiku, čo následne zvyšuje stav bdelosti a pripravenosť celého organizmu na ochranu.
Ochrana pred stresovými faktormi. Od práce Selyeho je známe, že celý hormonálny systém - predovšetkým hypofýza, ktorá sa nachádza v spodnej časti lebky, a kôra nadobličiek - sa aktivuje reťazovou reakciou, ktorá sa spúšťa zvýšenou aktivitou sympatiku. Preto je veľmi pravdepodobné, že celý komplexný systém obrany a adaptácie je regulovaný aktivujúcou retikulárnou formáciou. Škodlivé vonkajšie podnety (stresové účinky) aktivujú retikulárnu formáciu, a tým aj kôru, pozdĺž kolaterál zmyslových dráh. Zároveň sa zvyšuje aktivita sympatiku a uvoľňuje sa adrenalín, čím sa predlžuje bdelý stav kôry. Nešpecifický hormonálny obranný mechanizmus opísaný Cannonom a Selyem a retikulárny aktivačný systém sú teda funkčne prepojené.
Kontrola polohy tela. Retikulárny aktivačný systém sa podieľa nielen na hormonálnej regulácii, ale plní aj ďalšie funkcie. Jeho účasť na kontrole motorickej aktivity a udržiavaní svalového tonusu bola podrobne študovaná. Kôra a subkortikálne oblasti obsahujú množstvo špeciálnych motorických centier, ktoré tvoria súčasť komplexného samoregulačného motorického systému, ktorý zahŕňa mnoho častí od miechy až po kôru. V tomto systéme hrá dôležitú úlohu aj mozoček. Najnižšia - spinálna - úroveň eferentného motorického systému je pod kontrolou aktivačnej retikulárnej formácie. Toto riadenie sa uskutočňuje dvoma typmi vplyvov.
Po prvé, retikulárna formácia ovplyvňuje reflexy, ktoré kontrolujú polohu tela napriek gravitácii a iným vychyľovacím silám (tieto reflexy sme už spomínali v súvislosti s proprioceptormi, ktoré tvoria aferentný článok kortikálnej kontroly motorickej aktivity). Táto kontrola je pravdepodobne vykonávaná gama-eferentnými vláknami smerujúcimi do svalových vretien. Po druhé, retikulárne vplyvy hrajú dôležitú úlohu pri udržiavaní svalového tonusu v pokoji. Kostrové svaly sa neuvoľňujú, aj keď sú neaktívne, ale sú v stave určitého celkového tónu. Možno ich prirovnať k strunám klavíra, ktoré sa napínajú skôr, ako zaznejú nejaké zvuky. Aferentná aktivita proprioceptorov (svalových vretien) zohráva veľmi dôležitú úlohu v regulácii svalového tonusu (pozri str. 57). Takže
Retikulárna formácia teda pri obranných reakciách alebo pri agresii nielenže aktivuje kôru a spúšťa hormonálne reakcie, ale uvádza aj kostrové svalstvo do pohotovosti.
Regulácia činnosti autonómneho nervového systému. Už vyššie opísané funkcie naznačujú dôležitosť tejto relatívne malej skupiny neurónov. Okrem toho sú s retikulárnou formáciou spojené centrá životných funkcií nachádzajúce sa v predĺženej mieche: neuróny, ktoré riadia dýchanie, a jadrá, ktoré riadia krvný obeh a rôzne tráviace mechanizmy. Dá sa teda povedať, že difúzny retikulárny systém integruje všetku autonómnu aktivitu regulovanú mozgovým kmeňom.
Brzdné účinky. Ako už bolo spomenuté, stimuly, ktoré vyvolávajú impulznú aktivitu v extero- a interoceptoroch, aktivujú retikulárnu formáciu, ktorá následne zvyšuje aktivitu kôry. Niektoré aferentné impulzy však naopak retikulárnu aktivitu znižujú, a tým znižujú tonus kôry. Analýza vlákien IX a X hlavových nervov odhalila senzorické vlákna, ktoré vychádzajú zo steny aorty a krčnej tepny a ktorých stimulácia vedie k oslabeniu retikulárnej aktivity.
Fyziologická funkcia týchto inhibičných účinkov nie je úplne jasná. Možno tvoria súčasť inhibičného aparátu (Moruzzi a kol.), ktorý je zodpovedný za stav spánku. Inhibičné retikulárne neuróny možno nájsť vo vnútri aj mimo retikulárnej formácie. Tejto problematike sa budeme podrobne venovať v kapitole o spánku. Inhibičné mechanizmy sú tu uvedené len ako súčasť samoregulačného aparátu retikulárnej formácie.
Samoregulácia. Organizmus zvieraťa alebo človeka možno vzhľadom na jeho schopnosť samoregulácie považovať za automatický systém. Regulácia je v takomto systéme proces, ktorým sa udržiava stálosť určitých jeho parametrov. Pojem regulácie sa objavil v biológii dávno pred érou kybernetiky a počítačov. Okrem toho sa ako modely pre návrh automatických regulátorov použili rôzne biologické systémy. Princíp homeostázy opísaný Cannonom bol základom množstva zariadení určených na štúdium sebaregulácie.
Spätná väzba. Hlavným mechanizmom samoregulácie je spätná väzba, ktorá zabezpečuje nepretržitý tok informácií do riadiaceho centra o výsledkoch jeho príkazov. Účelom spätnej väzby je upraviť regulačné vplyvy, korigovať ich centrálnym regulačným aparátom. V technike sa spätná väzba využíva najmä
v servomechanizmoch. Automatické riadenie nie je možné bez spätnej väzby, teda bez toho, aby riadiace centrum dostávalo informácie o účinku jeho príkazov. Spätná väzba je pozitívna, ak vedie k zvýšeniu výslednej odchýlky, a negatívna, ak takúto odchýlku znižuje. Oba tieto spätnoväzbové mechanizmy fungujú v zmyslových systémoch a vo funkcii prebúdzania.
Centrum kontroly mozgu. Najdôležitejšou súčasťou všetkých samoregulačných automatických systémov je aparát, ktorý riadi prispôsobenie tela a zabezpečuje stálosť jeho parametrov. Táto časť sa nazýva riadiace centrum. Ľudský mozog možno považovať za centrum, kde mechanizmy spätnej väzby upravujú príkazy a tým zabezpečujú optimálne fungovanie.
Inhibícia retikulárnej formácie. Ak retikulárnu formáciu považujeme za blok regulujúci úroveň bdelosti tela, potom musí existovať mechanizmus spätnej väzby, ktorý by brzdil jeho prebúdzaciu aktivitu. Nedávno bolo opísaných niekoľko takýchto mechanizmov.
Kortikálna inhibícia. Prvým z týchto mechanizmov je inhibičný účinok kôry. Medzi kôrou a retikulárnou formáciou sú nielen vzostupné, ale aj zostupné vlákna, ktoré vedú impulzy z kôry. Takže retikulárne impulzy, ktoré regulujú tonus kôry, sú samy riadené kortikálnymi centrami. Tieto delenia spolu tvoria samoregulačný systém, vybudovaný na princípe reflexného krúžku.
Talamická inhibícia. Talamus obsahuje aj neurónovú sieť, ktorá sa pre svoju difúznu štruktúru nazývala talamická retikulárna formácia a ktorá sa podieľa na riadení retikulárnej aktivácie. Ako uvidíme, vysokofrekvenčné EEG s nízkou amplitúdou charakteristické pre bdelosť je výsledkom stimulácie retikulárnej formácie mozgového kmeňa a nízkofrekvenčná aktivita s vysokou amplitúdou počas spánku je spôsobená talamickou retikulárnou aktivitou. Talamické retikulárne bunky sú antagonistami retikulárnej formácie mozgového kmeňa. Elektrická stimulácia týchto neurónov talamu môže mať inhibičný účinok, ktorý spolu s ďalšími účinkami vedie k pomalej, synchronizovanej kortikálnej aktivite. Talamické inhibičné štruktúry prvýkrát opísal švajčiarsky fyziológ Hess asi pred tridsiatimi rokmi.
Medulárna inhibícia. Antagonisty retikulárnej aktivity sú tiež inhibičné neuróny v medulla oblongata, opísané Moruzzim. Aj keď sú tieto neuróny súčasťou retikulárnej formácie,

plnia nie aktivačnú, ale inhibičnú funkciu. Podľa Moruzziho a iných hrajú medulárne inhibičné neuróny dôležitú úlohu v mechanizme spánku.
Bdelosť mozgu je podporovaná samoregulačným systémom, ktorý pozostáva z retikulárnej formácie a vyššie popísaných inhibičných štruktúr. Úroveň kortikálneho tonusu, teda schopnosti mozgu byť stimulovaný zmyslovým vstupom, závisí od rovnováhy medzi aktivačnými a inhibičnými mechanizmami (obr. 38).



