Tanong47. Brainstem reticular formation
Ang konsepto ng neuroglia.
Neuroglia- ito ay mga cell na pumapalibot sa mga neuron at kasama sa kanila sa central nervous system at PNS. Ang bilang ng mga glial cell ay isang order ng magnitude na mas mataas kaysa sa bilang ng mga nerve cell.
Mga function ng neuroglia:
1.supporting - sumusuporta sa nerve cells
2.insulating - pinipigilan ang paglipat ng mga nerve impulses mula sa katawan ng isang neuron patungo sa katawan ng isa pa
3.regulatory - nakikilahok sa regulasyon ng central nervous system, lalo na, tinitiyak ang paghahatid ng mga impulses sa nais na direksyon
4. trophic - nakikilahok sa mga metabolic na proseso ng mga neuron
5. regulatory - kinokontrol ang excitability ng nerve cells.
Ang potensyal ng lamad (o potensyal na pahinga) ay ang potensyal na pagkakaiba sa pagitan ng panlabas at panloob na mga ibabaw ng lamad sa isang estado ng kamag-anak na physiological rest. Potensyal sa pagpapahinga bumangon bilang resulta ng dalawang dahilan:
1) hindi pantay na pamamahagi ng mga ion sa magkabilang panig ng lamad;
2) selective membrane permeability para sa mga ions. Sa pamamahinga, ang lamad ay hindi pareho natatagusan para sa iba't ibang mga ion. Ang lamad ng cell ay permeable sa K ions, mababa ang permeable sa Na ions at impermeable sa organic substance.
Dahil sa dalawang salik na ito, ang mga kondisyon ay nilikha para sa paggalaw ng mga ion. Ang paggalaw na ito ay isinasagawa nang walang pagkonsumo ng enerhiya sa pamamagitan ng passive transport - pagsasabog bilang isang resulta ng pagkakaiba sa konsentrasyon ng ion. Ang mga K ion ay umaalis sa selula at pinapataas ang positibong singil sa panlabas na ibabaw ng lamad; Ang mga Cl ions ay pasibo na pumapasok sa selula, na humahantong sa pagtaas ng positibong singil sa panlabas na ibabaw ng selula. Naiipon ang mga ion sa panlabas na ibabaw ng lamad at pinapataas ang positibong singil nito. Ang mga organikong compound ay nananatili sa loob ng cell. Bilang resulta ng paggalaw na ito, ang panlabas na ibabaw ng lamad ay positibong sinisingil, at ang panloob na ibabaw ay negatibong sisingilin. Ang panloob na ibabaw ng lamad ay maaaring hindi ganap na negatibong sisingilin, ngunit ito ay palaging negatibong sinisingil na may paggalang sa panlabas na isa. Ang estadong ito ng lamad ng cell ay tinatawag na estado ng polariseysyon. Ang paggalaw ng mga ion ay nagpapatuloy hanggang ang potensyal na pagkakaiba sa buong lamad ay balanse, ibig sabihin, ang electrochemical equilibrium ay nangyayari. Ang sandali ng balanse ay nakasalalay sa dalawang puwersa:
1) ang mga puwersa ng pagsasabog;
2) lakas pakikipag-ugnayan ng electrostatic. Electrochemical equilibrium value:
1) pagpapanatili ng ionic asymmetry;
2) pagpapanatili ng halaga ng potensyal ng lamad sa isang pare-parehong antas.
Ang puwersa ng pagsasabog (konsentrasyon pagkakaiba mga ion) at ang puwersa ng pakikipag-ugnayan ng electrostatic, samakatuwid ang potensyal ng lamad ay tinatawag na konsentrasyon-electrochemical.
Ang electrochemical equilibrium ay hindi sapat upang mapanatili ang ionic asymmetry. Sa isang hawla meron isa pang mekanismo ay ang sodium-potassium pump. Sodium-potassium pump - mekanismo tinitiyak ang aktibong transportasyon ng mga ion. Ang cell lamad ay naglalaman ng sistema carrier, na ang bawat isa ay nagbubuklod ng tatlong Na ion, na ay sa loob ng selda, at inilalabas sila. Mula sa labas, ang carrier ay nagbubuklod sa dalawang K ion sa labas ng cell at inililipat ang mga ito sa cytoplasm. Ang enerhiya ay kinuha mula sa pagkasira ng ATP.
2) (Ang mekanismo ng potensyal ng pahinga)
Ang potensyal na aksyon ay isang pagbabago sa potensyal ng lamad, umuusbong v mga tela sa ilalim ng pagkilos ng threshold at suprathreshold stimulus, na sinamahan ng recharging ng cell membrane.
Sa ilalim ng pagkilos ng threshold o suprathreshold stimulus, nagbabago ito pagkamatagusin cell lamad para sa mga ion sa iba't ibang antas. Para sa mga Na ions, tumataas ito at dahan-dahang umuunlad ang gradient. Bilang resulta, ang paggalaw ng mga Na ions ay nangyayari sa loob ng cell, mga ion Para umalis sa hawla na nangunguna upang muling magkarga ng lamad ng cell. Ang panlabas na ibabaw ng lamad ay nagdadala ng negatibong singil, ang panloob ay positibo.
Mga bahagi ng potensyal na pagkilos:
1) lokal na tugon;
2) mataas na boltahe peak potensyal (spike);
3) bakas ang pagbabagu-bago.
Ang mga Na ions ay pumapasok sa cell sa pamamagitan ng simpleng pagsasabog nang walang pagkonsumo ng enerhiya. Pag-abot sa threshold lakas, ang potensyal ng lamad ay bumababa sa isang kritikal na antas ng depolarization (mga 50 mV). Ang kritikal na antas ng depolarization ay ang bilang ng millivolts kung saan dapat bawasan ang potensyal ng lamad upang magkaroon ng mala-avalanche na kurso ng mga Na ions sa cell.
High voltage peak potential (spike).
Ang rurok ng potensyal na pagkilos ay isang palaging bahagi ng potensyal ng pagkilos. Binubuo ito ng dalawang yugto:
1) ang pataas na bahagi - ang yugto ng depolarization;
2) ang pababang bahagi - ang bahagi ng repolarization.
Ang pag-agos ng avalanche ng mga Na ions sa cell ay humahantong sa isang pagbabago sa potensyal sa lamad ng cell. Kapag mas maraming Na ions ang pumapasok sa cell, mas depolarized ang lamad, mas magbubukas ang activation gate. Ang hitsura ng isang singil na may kabaligtaran na tanda ay tinatawag na pagbabaligtad ng potensyal ng lamad. Ang paggalaw ng Na ions sa loob ng cell ay nagpapatuloy hanggang sa sandali ng electrochemical equilibrium na may paggalang sa Na ion. Ang amplitude ng action potential ay hindi nakasalalay sa lakas ng stimulus, depende ito sa konsentrasyon ng Na ions at sa antas ng lamad pagkamatagusin sa Na ions. Ang pababang bahagi (repolarization phase) ay nagbabalik ng lamad na singil sa orihinal nitong tanda. Sa pag-abot sa electrochemical equilibrium para sa Na ions, ang activation gate ay inactivated, pagkamatagusin sa Na ions at ang permeability sa K ions ay tumataas. Ang kumpletong pagpapanumbalik ng potensyal ng lamad ay hindi mangyayari.
Sa proseso ng pagbawi mga reaksyon Ang mga potensyal na bakas ay naitala sa lamad ng cell - positibo at negatibo.
3) (Pagbabago sa excitability kapag pumasa sa isang alon ng kaguluhan)
Sa pag-unlad ng potensyal na pagkilos, ang isang pagbabago sa excitability ng tissue ay nangyayari, at ang pagbabagong ito ay nagpapatuloy sa mga yugto (Larawan 2). Ang estado ng paunang polariseysyon ng lamad, na sinasalamin ng potensyal ng resting lamad, ay tumutugma sa paunang estado ng excitability nito at, samakatuwid, ang mga cell ay ang normal na antas ng excitability. Sa panahon ng pre-soldering, ang tissue excitability ay nadagdagan, ang yugtong ito ng excitability ay tinatawag pangunahing kadakilaan. Sa panahon ng pagbuo ng pre-soldering, ang resting membrane potential ay lumalapit sa kritikal na antas ng depolarization, at upang makamit ang huli, ang lakas ng stimulus, na mas mababa sa threshold (subthreshold), ay sapat.
Sa panahon ng pag-unlad ng spike (peak potential), ang isang mala-avalanche na daloy ng sodium ions sa cell ay nangyayari, bilang isang resulta kung saan ang lamad ay muling na-recharge at nawawala ang kakayahang tumugon nang may paggulo sa stimuli kahit na sa isang suprathreshold. puwersa. Ang yugtong ito ng excitability ay tinatawag ganap na refractoriness(ganap na di-excitability). Ito ay tumatagal hanggang sa katapusan ng recharge ng lamad. Ang ganap na refractoriness, iyon ay, kumpletong non-excitability ng lamad, ay lumitaw dahil sa ang katunayan na ang mga channel ng sodium ay ganap na binuksan sa simula at pagkatapos ay hindi aktibo.
Matapos ang pagtatapos ng yugto ng recharge ng lamad, ang excitability nito ay unti-unting naibalik sa orihinal na antas nito - yugto ng relatibong refractoriness. Nagpapatuloy ito hanggang sa maibalik ang singil ng lamad sa isang halaga na tumutugma sa kritikal na antas ng depolarization. Dahil sa panahong ito ang potensyal ng lamad ng pahinga ay hindi pa naibabalik, ang excitability ng tissue ay nabawasan at ang bagong paggulo ay maaaring lumitaw lamang sa ilalim ng pagkilos ng isang suprathreshold stimulus. Ang pagbawas sa excitability sa yugto ng kamag-anak na refractoriness ay nauugnay sa bahagyang hindi aktibo ng mga channel ng sodium at pag-activate ng mga channel ng potassium.
Ang panahon ng negatibong bakas na potensyal ay tumutugma sa isang mas mataas na antas ng excitability - yugto ng pangalawang kadakilaan. Dahil ang potensyal ng lamad sa yugtong ito ay mas malapit sa kritikal na antas ng depolarization, ngunit sa paghahambing sa estado ng pahinga (paunang polariseysyon), ang threshold ng pangangati ay nabawasan, ibig sabihin, ang excitability ay nadagdagan. Sa yugtong ito, maaaring lumitaw ang bagong kaguluhan sa ilalim ng pagkilos ng stimuli ng subthreshold force. Ang mga channel ng sodium ay hindi ganap na inactivate sa yugtong ito. Sa panahon ng pagbuo ng isang positibong bakas na potensyal, ang excitability ng tissue ay nabawasan - yugto ng pangalawang refractoriness. Sa yugtong ito, tumataas ang potensyal ng lamad (ang estado ng hyperpolarization ng lamad), lumalayo mula sa kritikal na antas ng depolarization, tumataas ang threshold ng pangangati at ang bagong paggulo ay maaaring lumitaw lamang sa ilalim ng pagkilos ng stimuli ng isang halaga ng suprathreshold. Ang hyperpolarization ng lamad ay bubuo dahil sa tatlong dahilan: una, ang patuloy na paglabas ng mga potassium ions; pangalawa, sa pamamagitan ng pagbubukas, posibleng, ng mga channel para sa chlorine at ang pagpasok ng mga ion na ito sa cytoplasm ng cell; pangatlo, ang pinahusay na gawain ng sodium-potassium pump.
4) (Nagsasagawa ng paggulo kasama ang mga nerve fibers)
Ang mekanismo ng pagpapalaganap ng paggulo sa iba't ibang mga nerve fibers ay hindi pareho. Ayon sa mga modernong konsepto, ang pagpapalaganap ng paggulo kasama ang mga nerve fibers ay isinasagawa batay sa mga mekanismo ng ionic para sa pagbuo ng isang potensyal na aksyon.
Sa pagpapalaganap ng excitation sa kahabaan ng myelin-free nerve fiber, ang mga lokal na electric current na lumabas sa pagitan ng nasasabik na seksyon nito, negatibong sisingilin, at hindi nasasabik, positibong sisingilin, ay nagdudulot ng depolarization ng lamad sa kritikal na antas, na sinusundan ng pagbuo ng AP sa pinakamalapit na punto. ng hindi nasasabik na seksyon ng lamad. Ang prosesong ito ay paulit-ulit nang maraming beses. Sa buong haba ng nerve fiber, ang proseso ng pagpaparami ng bagong AP ay nangyayari sa bawat punto ng fiber membrane. Tinatawag mo itong pag-uugali ng pagpukaw at tuloy-tuloy.
Ang pagkakaroon ng isang kaluban na may mataas na resistensya ng kuryente sa mga hibla ng myelin, pati na rin ang mga seksyon ng hibla na walang kaluban (mga pagharang ni Ranvier) lumikha ng mga kondisyon para sa isang qualitatively bagong uri ng pagpapadaloy ng paggulo kasama ang myelinated nerve fibers. Lumilitaw ang mga lokal na agos ng kuryente sa pagitan ng mga katabing interception ng Ranvier, dahil ang nasasabik na intercept membrane ay nagiging negatibong sisingilin kaugnay sa ibabaw ng katabing unexcited intercept. Ang mga lokal na alon na ito ay nagde-depolarize ng lamad ng hindi nasasabik na pagharang sa isang kritikal na antas at ang AP ay lilitaw dito (Larawan 4). Dahil dito, ang kaguluhan ay tila "tumalon" sa mga lugar ng nerve fiber, na sakop ng myelin, mula sa isang pagharang patungo sa isa pa. Ang ganitong mekanismo para sa pagpapalaganap ng paggulo ay tinatawag maalat o palpak. Ang bilis ng pamamaraang ito ng pagsasagawa ng paggulo ay mas mataas at ito ay mas matipid kumpara sa patuloy na pagsasagawa ng paggulo, dahil hindi ang buong lamad ay kasangkot sa estado ng aktibidad, ngunit ang maliliit na seksyon lamang nito sa lugar ng mga interceptions.
kanin. 4. Scheme ng pagpapalaganap ng excitation sa myelin-free (a) at myelinic (b) nerve fibers.
Ang "paglukso" ng potensyal na pagkilos sa ibabaw ng seksyon sa pagitan ng mga interception ay posible dahil ang AP amplitude ay 5-6 beses na mas mataas kaysa sa halaga ng threshold na kinakailangan upang ma-excite ang isang kalapit na interception. Ang PD ay maaaring "tumalon" hindi lamang sa pamamagitan ng isa, kundi pati na rin sa pamamagitan ng dalawang inter-intercept gaps. Ang hindi pangkaraniwang bagay na ito ay maaaring maobserbahan sa isang pagbawas sa excitability ng isang kalapit na interception sa ilalim ng impluwensya ng anumang pharmacological substance, halimbawa, novocaine, cocaine, atbp.
Ang mga hibla ng nerbiyos ay nagtataglay lability- ang kakayahang magparami ng isang tiyak na bilang ng mga siklo ng paggulo sa bawat yunit ng oras alinsunod sa ritmo ng kumikilos na stimuli. Ang sukatan ng lability ay ang maximum na bilang ng mga siklo ng paggulo na maaaring kopyahin ng isang nerve fiber bawat yunit ng oras nang walang pagbabago sa ritmo ng pagpapasigla. Ang lability ay tinutukoy ng tagal ng rurok ng potensyal ng pagkilos, iyon ay, sa pamamagitan ng yugto ng ganap na refractoriness. Dahil ang tagal ng absolute refraction sa spike potential ng nerve fiber ay ang pinakamaikling, ang lability nito ay ang pinakamataas. Ang nerve fiber ay may kakayahang magparami ng hanggang 1000 impulses bawat segundo.
Natuklasan ni N.E. Vvedensky na kung malantad ang isang seksyon ng nerve mga pagbabago(ibig sabihin, pagkakalantad sa isang nakakapinsalang ahente) sa pamamagitan ng, halimbawa, pagkalason o pinsala, pagkatapos ay ang lability ng naturang site ay nabawasan nang husto. Ang pagpapanumbalik ng paunang estado ng nerve fiber pagkatapos ng bawat potensyal na pagkilos sa nasirang lugar ay nangyayari nang dahan-dahan. Kapag ang site na ito ay nalantad sa madalas na stimuli, hindi nito magagawang kopyahin ang ibinigay na ritmo ng pagpapasigla, at samakatuwid ang pagpapadaloy ng mga impulses ay naharang. Ang estado ng pinababang lability na ito ay pinangalanan ni N.E. Vvedensky parabiosis. Sa pag-unlad ng estado ng parabiosis, tatlong sunud-sunod na pagpapalit ng mga phase ay maaaring mapansin: equalizing, paradoxical, inhibitory.
V yugto ng equalizing mayroong pagkakapantay-pantay ng magnitude ng pagtugon sa madalas at pambihirang stimuli. Sa ilalim ng normal na mga kondisyon ng paggana ng nerve fiber, ang magnitude ng tugon ng mga fibers ng kalamnan na innervated nito ay sumusunod sa batas ng puwersa: ang tugon sa bihirang stimuli ay mas mababa, at sa madalas na stimuli - higit pa. Sa ilalim ng pagkilos ng isang ahente ng parabiotic at may isang bihirang ritmo ng pangangati (halimbawa, 25 Hz), ang lahat ng mga impulses ng paggulo ay isinasagawa sa pamamagitan ng site ng parabiotic, dahil ang excitability pagkatapos ng nakaraang impulse ay may oras upang mabawi. Sa mataas na ritmo ng pagpapasigla (100 Hz), ang mga kasunod na impulses ay maaaring dumating sa oras na ang nerve fiber ay nasa estado pa rin ng relatibong refractoriness na dulot ng nakaraang potensyal na pagkilos. Samakatuwid, ang ilan sa mga impulses ay hindi isinasagawa. Kung ang bawat ika-apat na paggulo ay isinasagawa lamang (iyon ay, 25 impulses mula sa 100), kung gayon ang amplitude ng tugon ay magiging pareho sa bihirang stimuli (25 Hz) - nangyayari ang isang pagkakapantay-pantay ng tugon.
V kabalintunaan-phase mayroong karagdagang pagbaba sa lability. Kasabay nito, ang isang tugon ay nangyayari sa bihira at madalas na stimuli, ngunit ito ay mas kaunti sa madalas na stimuli, dahil ang madalas na stimuli ay higit na nagpapababa ng lability, na nagpapahaba sa yugto ng absolute refractoriness. Dahil dito, mayroong isang kabalintunaan - ang tugon sa mga bihirang stimuli ay mas malaki kaysa sa madalas.
V yugto ng pagpepreno ang lability ay bumababa sa isang lawak na ang parehong bihira at madalas na stimuli ay hindi nagiging sanhi ng isang tugon. Sa kasong ito, ang lamad ng nerve fiber ay depolarized at hindi napupunta sa yugto ng repolarization, iyon ay, ang orihinal na estado nito ay hindi naibalik.
Ang kababalaghan ng parabiosis ay ang batayan ng lokal na kawalan ng pakiramdam na dulot ng droga. Ang impluwensya ng anesthetics ay nauugnay din sa isang pagbawas sa lability at isang paglabag sa mekanismo ng pagpapadaloy ng paggulo kasama ang mga nerve fibers.
Ang parabiosis ay isang nababaligtad na kababalaghan. Kung ang parabiotic substance ay kumikilos sa loob ng maikling panahon, pagkatapos pagkatapos ng pagwawakas ng pagkilos nito, ang nerve ay umalis sa estado ng parabiosis sa pamamagitan ng parehong mga yugto, ngunit sa kabaligtaran na pagkakasunud-sunod.
Ang mekanismo ng pag-unlad ng estado ng parabiotic ay ang mga sumusunod. Kapag ang isang parabiotic factor ay kumikilos sa isang nerve fiber, ang kakayahan ng lamad na pataasin ang sodium permeability bilang tugon sa pangangati ay naaabala. Sa lugar ng pagbabago, ang hindi aktibo ng mga channel ng sodium na dulot ng isang nakakapinsalang ahente ay idinagdag sa hindi aktibo na dulot ng isang nerve impulse, at ang excitability ay bumaba nang labis na ang pagpapadaloy ng susunod na salpok ay naharang.
5) (Synapses, ang kanilang mga uri, mga tampok sa istruktura)
Physiology ng synapses.
Sa CNS, ang mga nerve cell ay konektado sa isa't isa sa pamamagitan ng synapses. Synapse Ay isang structurally functional formation na nagsisiguro sa paghahatid ng excitation o inhibition mula sa isang nerve fiber patungo sa isang innervated cell.
Synapses sa lokalisasyon ay nahahati sa gitnang (matatagpuan sa loob ng central nervous system, pati na rin sa ganglia ng autonomic nervous system) at peripheral (na matatagpuan sa labas ng central nervous system, nagbibigay ng komunikasyon sa mga cell ng innervated tissue).
Functionally Ang mga synapses ay nahahati sa kapana-panabik, kung saan, bilang resulta ng depolarization ng postsynaptic membrane, nabuo ang isang excitatory postsynaptic na potensyal, at preno, sa mga presynaptic na dulo kung saan ang isang tagapamagitan ay inilabas, na naghi-hyperpolarize ng postsynaptic membrane at nagiging sanhi ng paglitaw ng isang nagbabawal na potensyal na postsynaptic.
Sa pamamagitan ng mekanismo ng paghahatid Ang mga synapses ay nahahati sa kemikal at elektrikal. Ang mga synapses ng kemikal ay nagpapadala ng kaguluhan o pagsugpo dahil sa mga espesyal na sangkap - mga tagapamagitan. Depende sa mula sa uri ng tagapamagitan chemical synapses ay nahahati sa:
1.cholinergic (tagapamagitan - acetylcholine)
2.adrenergic (mga tagapamagitan - adrenaline, norepinephrine)
Anatomical na pag-uuri Ang mga synapses ay nahahati sa neurosecretory, neuromuscular at interneuronal.
Synapse ay binubuo ng tatlong pangunahing sangkap:
1.presynaptic membrane
2. Postsynaptic lamad
3.synaptic cleft
Ang presynaptic membrane ay ang dulo ng proseso ng nerve cell. Sa loob ng apendiks, sa agarang paligid ng lamad, mayroong isang akumulasyon ng mga vesicle (mga butil) na naglalaman ng isa o ibang tagapamagitan. Ang mga bula ay patuloy na gumagalaw.
Ang postsynaptic membrane ay bahagi ng innervated tissue cell membrane. Ang postsynaptic membrane, sa kaibahan sa presynaptic membrane, ay may mga chemoreceptor ng protina sa biologically active (mediators, hormones), medicinal at toxic substances. Ang isang mahalagang katangian ng mga postsynaptic membrane receptor ay ang kanilang chemical specificity, i.e. ang kakayahang pumasok sa biochemical interaction lamang sa isang tiyak na uri ng tagapamagitan.
Ang synaptic cleft ay ang espasyo sa pagitan ng pre- at postsynaptic membranes, na puno ng likidong katulad ng komposisyon sa plasma ng dugo. Sa pamamagitan nito, ang tagapamagitan ay dahan-dahang nagkakalat mula sa presynaptic membrane hanggang sa postsynaptic.
Ang mga tampok na istruktura ng neuromuscular synapse ay tumutukoy nito pisyolohikal na katangian.
1. Unilateral conduction ng excitation (mula sa pre- to the postsynaptic membrane), dahil sa pagkakaroon ng mga receptor na sensitibo sa mediator lamang sa postsynaptic membrane.
2. Synaptic delay sa conduction ng excitation (ang oras sa pagitan ng pagdating ng impulse sa presynaptic end at ang simula ng postsynaptic response), na nauugnay sa mababang diffusion rate ng transmitter sa synaptic cleft kumpara sa bilis ng ang salpok sa pamamagitan ng nerve fiber.
3. Mababang lability at mataas na pagkapagod ng synapse dahil sa oras ng pagpapalaganap ng nakaraang salpok at ang pagkakaroon ng isang panahon ng ganap na refractoriness sa loob nito.
4. Mataas na selective sensitivity ng synapse sa mga kemikal, dahil sa pagiging tiyak ng mga chemoreceptor ng postsynaptic membrane.
Mga yugto ng synaptic transmission.
1. Synthesis ng isang tagapamagitan... Sa cytoplasm ng mga neuron at nerve endings, ang mga chemical mediator ay synthesized - biologically active substances. Ang mga ito ay patuloy na synthesize at idineposito sa synaptic vesicle ng nerve endings.
2. Ang pagtatago ng transmiter... Ang paglabas ng isang neurotransmitter mula sa synaptic vesicle ay isang quantum na kalikasan. Sa pamamahinga, ito ay hindi gaanong mahalaga, ngunit sa ilalim ng impluwensya ng isang nerve impulse ay tumataas ito nang husto.
3. Pakikipag-ugnayan ng isang tagapamagitan sa mga receptor ng postsynaptic membrane... Ang pakikipag-ugnayan na ito ay binubuo sa piling pagbabago ng permeability ng mga ion-selective na channel ng effector cell sa rehiyon ng mga aktibong sentro ng pagbubuklod sa mediator. Ang pakikipag-ugnayan ng isang tagapamagitan sa mga receptor nito ay maaaring maging sanhi ng paggulo o pagsugpo ng isang neuron, pag-urong ng isang selula ng kalamnan, ang pagbuo at pagpapalabas ng mga hormone sa pamamagitan ng mga selulang nagtatago. Sa kaso ng isang pagtaas sa pagkamatagusin ng sodium at calcium channels, ang pagpasok ng Na at Ca sa cell ay tumataas, na sinusundan ng depolarization ng lamad, ang hitsura ng PD at karagdagang paghahatid ng nerve impulse. Ang ganitong mga synapses ay tinatawag na excitatory. Kung ang pagkamatagusin ng mga channel ng potassium at mga channel para sa chlorine ay tumaas, mayroong isang labis na pagpapalabas ng K mula sa cell na may sabay-sabay na pagsasabog ng Cl dito, na humahantong sa hyperpolarization ng lamad, isang pagbawas sa excitability nito at ang pagbuo ng mga nagbabawal na mga potensyal na postsynaptic. Ang paghahatid ng mga nerve impulses ay nagiging mahirap o ganap na huminto. Ang ganitong mga synapses ay tinatawag na inhibitory synapses.
Ang mga receptor na nakikipag-ugnayan sa ACh ay tinatawag na mga cholinergic receptor. Sa paggana, nahahati sila sa dalawang grupo: M - at H-cholinergic receptors. Sa mga synapses ng mga kalamnan ng kalansay, ang mga H-cholinergic receptor lamang ang naroroon, habang sa mga kalamnan ng mga panloob na organo, pangunahin ang mga M-cholinergic receptor na naroroon.
Ang mga receptor na nakikipag-ugnayan sa HA ay tinatawag na mga adrenergic receptor. Sa paggana, nahahati sila sa alpha at beta adrenergic receptors. Sa postsynaptic membrane ng makinis na mga selula ng kalamnan ng mga panloob na organo at mga daluyan ng dugo, ang parehong uri ng mga adrenergic receptor ay madalas na magkakasamang nabubuhay. Ang pagkilos ng NA ay depolarizing kung ito ay nakikipag-ugnayan sa mga alpha-adrenergic receptors (contraction ng muscular membrane ng mga pader ng mga daluyan ng dugo o bituka), o nakakapigil - kapag ito ay nakikipag-ugnayan sa mga beta-adrenergic receptors (ang kanilang pagpapahinga).
4. Hindi aktibo ang transmiter... Ang hindi aktibo (kumpletong pagkawala ng aktibidad) ng tagapamagitan ay kinakailangan para sa repolarization ng postsynaptic membrane at pagpapanumbalik ng paunang antas ng MP. Ang pinakamahalagang ruta ng hindi aktibo ay hydrolytic degradation na may mga inhibitor. Para sa ACh, ang inhibitor ay cholinesterase, para sa HA at adrenaline - monoamine oxidase at catecholoxymethyltransferase.
Ang isa pang paraan ng pag-alis ng isang tagapamagitan mula sa synaptic cleft ay ang "reuptake" nito sa pamamagitan ng presynaptic endings nito (pinocytosis) at reverse axonal transport, lalo na binibigkas para sa mga catecholamines.
Ang aktibidad ng koordinasyon ng central nervous system ay batay sa pakikipag-ugnayan ng mga proseso ng paggulo at pagsugpo.
Excitation Ay isang aktibong proseso na isang tugon ng tissue sa pangangati at nailalarawan sa pamamagitan ng pagtaas ng mga function ng tissue.
Pagpreno Ay isang aktibong proseso na isang tugon ng tissue sa pangangati at nailalarawan sa pamamagitan ng pagbaba sa mga function ng tissue.
Ang pangunahing pagsugpo sa central nervous system ay nangyayari dahil sa mga inhibitory neuron. Ito ay isang espesyal na uri ng mga intercalary neuron na, kapag nagpapadala ng isang salpok, naglalabas ng isang nagbabawal na neurotransmitter. Mayroong 2 uri ng pangunahing pagsugpo: postsynaptic at presynaptic.
Postsynaptic inhibition ay nangyayari kapag ang axon ng isang inhibitory neuron ay bumubuo ng isang synapse sa katawan ng neuron at, naglalabas ng isang transmiter, nagiging sanhi ng hyperpolarization ng cell lamad, inhibiting ang aktibidad ng cell.
Presynaptic inhibition nangyayari kapag ang axon ng inhibitory neuron ay bumubuo ng isang synapse na may axon ng excitatory neuron, na pumipigil sa pagpapadaloy ng salpok.
6) (Spinal cord, mga pag-andar nito, pakikilahok sa regulasyon ng tono ng kalamnan)
Ang spinal cord ay gumaganap ng reflex at conduction function. Ang una ay ibinibigay ng mga sentro ng nerbiyos nito, ang pangalawa sa pamamagitan ng mga landas.
Mayroon itong segmental na istraktura. Bukod dito, ang paghahati sa mga segment ay gumagana. Ang bawat segment ay bumubuo ng anterior at posterior roots. Ang mga hulihan ay sensitibo, i.e. afferent, anterior motor, efferent. Ang pattern na ito ay tinatawag na batas ni Bella-Magendie. Ang mga ugat ng bawat segment ay nagpapaloob sa 3 body metameres, ngunit bilang resulta ng overlapping, ang bawat metamere ay innervate ng tatlong segment. Samakatuwid, kapag ang mga nauunang ugat ng isang segment ay apektado, ang aktibidad ng motor ng kaukulang metamer ay humina lamang.
Morphologically, ang mga katawan ng mga neuron ng spinal cord ay bumubuo ng kulay abong bagay nito. Sa paggana, ang lahat ng mga neuron nito ay nahahati sa mga motoneuron, intercalary, mga neuron ng nagkakasundo at parasympathetic na mga dibisyon ng autonomic nervous system. Ang mga motor neuron, depende sa kanilang functional significance, ay nahahati sa alpha at gamma motor neuron. Ang mga hibla ng afferent pathway ay napupunta sa a-motoneurons, na nagsisimula sa intrafusal, i.e. receptor na mga selula ng kalamnan. Ang mga katawan ng a-motor neuron ay matatagpuan sa mga anterior horn ng spinal cord, at ang kanilang mga axon ay nagpapapasok sa mga kalamnan ng kalansay. Ang mga gamma motoneuron ay kinokontrol ang tensyon ng spindle ng kalamnan i.e. intrafusal fibers. Kaya, sila ay kasangkot sa regulasyon ng mga contraction ng skeletal muscle. Samakatuwid, kapag ang mga nauunang ugat ay pinutol, ang tono ng kalamnan ay nawawala.
Ang mga interneuron ay nagbibigay ng komunikasyon sa pagitan ng mga sentro ng spinal cord at ang mga nakapatong na bahagi ng central nervous system.
Ang mga neuron ng nagkakasundo na bahagi ng autonomic nervous system ay matatagpuan sa mga lateral horns ng thoracic segment, at ang parasympathetic sa sacral na bahagi.
Ang conductive function ay upang matiyak ang koneksyon ng peripheral receptors, mga sentro ng spinal cord sa mga nakapatong na bahagi ng central nervous system, pati na rin ang mga nerve center nito sa isa't isa. Ito ay isinasagawa sa pamamagitan ng mga landas. Ang lahat ng mga pathway ng spinal cord ay nahahati sa sarili o propriospinal, pataas at pababa. Ang mga propriospinal pathway ay nagkokonekta sa mga nerve center ng iba't ibang mga segment ng spinal cord. Ang kanilang pag-andar ay upang i-coordinate ang tono ng kalamnan, mga paggalaw ng iba't ibang mga metamer ng puno ng kahoy.
Ang mga pataas na landas ay kinabibilangan ng ilang mga landas. Ang mga bundle ni Gaulle at Burdach ay nagsasagawa ng mga nerve impulses mula sa proprioceptors ng mga kalamnan at tendon hanggang sa kaukulang nuclei ng medulla oblongata, at pagkatapos ay sa thalamus at somatosensory area ng cortex. Salamat sa mga landas na ito, ang postura ng katawan ay tinasa at naitama. Ang Govers at Fleksig bundle ay nagpapadala ng paggulo mula sa proprioceptors, mechanoreceptors ng balat hanggang sa cerebellum. Dahil dito, natiyak ang pang-unawa at walang malay na koordinasyon ng pustura. Ang mga spinothalamic tract ay nagsasagawa ng mga senyales mula sa sakit, temperatura, mga tactile receptor ng balat hanggang sa thalamus, at pagkatapos ay ang mga somatosensory zone ng cortex. Tinitiyak nila ang pang-unawa ng kaukulang mga signal at ang pagbuo ng sensitivity.
Ang mga pababang landas ay nabuo din ng ilang mga landas. Ang mga corticospinal pathway ay napupunta mula sa pyramidal at extrapyramidal neurons ng cortex hanggang sa a-motor neurons ng spinal cord. Dahil sa kanila, ang regulasyon ng mga boluntaryong paggalaw ay isinasagawa. Ang rubrospinal pathway ay nagsasagawa ng mga signal mula sa pulang nucleus ng midbrain hanggang sa mga a-motor neuron ng mga flexor na kalamnan. Ang vestibulospinal pathway ay nagpapadala ng mga signal mula sa vestibular nuclei ng medulla oblongata, pangunahin ang Deiters nucleus, sa mga a-motor neuron ng mga extensor na kalamnan. Dahil sa dalawang landas na ito, ang tono ng kaukulang mga kalamnan ay kinokontrol sa mga pagbabago sa posisyon ng katawan.
Ang lahat ng mga reflexes ng spinal cord ay nahahati sa somatic, i.e. motor at vegetative. Ang mga somatic reflexes ay nahahati sa tendon o myotatic at cutaneous. Ang mga tendon reflexes ay nangyayari sa mekanikal na pangangati ng mga kalamnan at tendon. Ang kanilang bahagyang pag-uunat ay humahantong sa paggulo ng mga tendon receptor at a-motoneuron ng spinal cord. Bilang isang resulta, ang pag-urong ng kalamnan ay nangyayari, lalo na ang mga extensor. Kasama sa mga tendon reflexes ang tuhod, Achilles, siko, pulso, atbp., na nagmumula sa mekanikal na pangangati ng kaukulang mga tendon. Halimbawa, ang tuhod ay ang pinakasimpleng monosynaptic, dahil mayroon lamang isang synapse sa gitnang bahagi nito. Ang mga reflexes ng balat ay sanhi ng pangangati ng mga receptor ng balat, ngunit ipinakikita ng mga reaksyon ng motor. Ang mga ito ay plantar at abdominal (paliwanag). Ang mga spinal nerve center ay nasa ilalim ng kontrol ng mga nasa ibabaw. Samakatuwid, pagkatapos ng seksyon sa pagitan ng medulla oblongata at ng spinal cord, nangyayari ang pagkabigla ng gulugod at ang tono ng lahat ng mga kalamnan ay makabuluhang bababa.
Ang mga autonomic reflexes ng spinal cord ay nahahati sa sympathetic at parasympathetic. Parehong ipinakita ang reaksyon ng mga panloob na organo sa pangangati ng mga receptor ng balat, mga panloob na organo, mga kalamnan. Ang mga vegetative neuron ng spinal cord ay bumubuo sa mas mababang mga sentro para sa regulasyon ng vascular tone, aktibidad ng puso, bronchial lumen, pagpapawis, pag-ihi, pagdumi, pagtayo, bulalas, atbp.
7) (Ang medulla oblongata at pons, ang kanilang mga pag-andar, pakikilahok sa regulasyon ng tono ng kalamnan)
Medulla
Mga tampok ng functional na organisasyon. Ang medulla oblongata (medullaoblongata) sa mga tao ay may haba na humigit-kumulang 25 mm. Ito ay extension ng spinal cord. Sa istruktura, sa mga tuntunin ng pagkakaiba-iba at istraktura ng nuclei, ang medulla oblongata ay mas kumplikado kaysa sa spinal cord. Hindi tulad ng spinal cord, wala itong metameric, paulit-ulit na istraktura; ang kulay abong bagay sa loob nito ay hindi matatagpuan sa gitna, ngunit sa nuclei patungo sa periphery.
Sa medulla oblongata, may mga olibo na nauugnay sa spinal cord, extrapyramidal system at cerebellum - ito ay manipis at hugis-wedge na nuclei ng proprioceptive sensitivity (Gaulle at Burdach nuclei). Narito ang mga intersection ng pababang pyramidal path at ang pataas na mga landas na nabuo ng manipis at hugis-wedge na mga bundle (Gaulle at Burdach), ang reticular formation.
Ang medulla oblongata, dahil sa mga nuclear formations at reticular formation nito, ay nakikilahok sa pagpapatupad ng vegetative, somatic, gustatory, auditory, vestibular reflexes. Ang isang tampok ng medulla oblongata ay ang nuclei nito, na nasasabik nang sunud-sunod, ay nagbibigay ng pagpapatupad ng mga kumplikadong reflexes na nangangailangan ng sunud-sunod na pag-activate ng iba't ibang mga grupo ng kalamnan, na sinusunod, halimbawa, kapag lumulunok.
Ang nuclei ng mga sumusunod na cranial nerves ay matatagpuan sa medulla oblongata:
isang pares ng VIII cranial nerves - ang vestibular cochlear nerve ay binubuo ng mga bahagi ng cochlear at vestibular. Ang cochlear nucleus ay nasa medulla oblongata;
pares IX - glossopharyngeal nerve (item glossopharyngeus); ang core nito ay nabuo ng 3 bahagi - motor, sensory at vegetative. Ang bahagi ng motor ay nakikilahok sa innervation ng mga kalamnan ng pharynx at oral cavity, ang sensitibong bahagi ay tumatanggap ng impormasyon mula sa mga receptor ng lasa ng posterior third ng dila; vegetative innervates ang salivary glands;
pares X - ang vagus nerve (n.vagus) ay may 3 nuclei: ang vegetative ay nagpapapasok ng larynx, esophagus, puso, tiyan, bituka, digestive glands; ang sensitibo ay tumatanggap ng impormasyon mula sa mga receptor ng alveoli ng mga baga at iba pang mga panloob na organo at ang motor (ang tinatawag na mutual) ay nagbibigay ng isang pagkakasunud-sunod ng pag-urong ng mga kalamnan ng pharynx at larynx kapag lumulunok;
pares XI - accessory nerve (n.accessorius); ang nucleus nito ay bahagyang matatagpuan sa medulla oblongata;
pares XII - ang hypoglossal nerve (n.hypoglossus) ay ang motor nerve ng dila, ang nucleus nito ay kadalasang matatagpuan sa medulla oblongata.
Mga function ng pandama. Kinokontrol ng medulla oblongata ang isang bilang ng mga sensory function: ang pagtanggap ng sensitivity ng balat ng mukha - sa sensory nucleus ng trigeminal nerve; pangunahing pagsusuri ng pagtanggap ng lasa - sa nucleus ng glossopharyngeal nerve; pagtanggap ng auditory stimuli - sa nucleus ng cochlear nerve; pagtanggap ng vestibular stimuli - sa itaas na vestibular nucleus. Sa posterior superior na bahagi ng medulla oblongata, may mga daanan ng cutaneous, deep, visceral sensitivity, ang ilan sa mga ito ay inililipat dito sa pangalawang neuron (manipis at hugis-wedge na nuclei). Sa antas ng medulla oblongata, ang mga sensory function sa itaas ay nagpapatupad ng pangunahing pagsusuri ng lakas at kalidad ng pagpapasigla, pagkatapos ang naprosesong impormasyon ay ipinadala sa mga subcortical na istruktura upang matukoy ang biological na kahalagahan ng pagpapasigla na ito.
Pagsasagawa ng mga function. Ang lahat ng pataas at pababang mga landas ng spinal cord ay dumadaan sa medulla oblongata: spinal thalamic, corticospinal, rubrospinal. Ito ang pinagmulan ng vestibulospinal, olivospinal at reticulospinal tracts, na nagbibigay ng tono at koordinasyon ng mga reaksyon ng kalamnan. Sa medulla oblongata, ang mga landas mula sa cerebral cortex ay nagtatapos - ang mga cortex-reticular na landas. Narito ang mga pataas na landas ng proprioceptive sensitivity mula sa dulo ng spinal cord: manipis at hugis-wedge. Ang mga pormasyon ng utak tulad ng tulay, midbrain, cerebellum, thalamus, hypothalamus at cerebral cortex ay may dalawang-daan na koneksyon sa medulla oblongata. Ang pagkakaroon ng mga koneksyon na ito ay nagpapahiwatig ng pakikilahok ng medulla oblongata sa regulasyon ng tono ng kalamnan ng kalansay, autonomic at mas mataas na integrative function, at ang pagsusuri ng sensory stimuli.
Mga function ng reflex. Maraming reflexes ng medulla oblongata ay nahahati sa vital at non-vital, gayunpaman, ang konsepto na ito ay sa halip arbitrary. Ang mga sentro ng paghinga at vasomotor ng medulla oblongata ay maaaring maiugnay sa mga mahahalagang sentro, dahil ang isang bilang ng mga cardiac at respiratory reflexes ay sarado sa kanila.
Ang medulla oblongata ay nag-aayos at nagpapatupad ng isang bilang ng mga proteksiyon na reflexes: pagsusuka, pagbahin, pag-ubo, pagpunit, pagsasara ng mga talukap ng mata. Ang mga reflexes na ito ay natanto dahil sa ang katunayan na ang impormasyon tungkol sa pangangati ng mga receptor ng mucous membrane ng mata, bibig, larynx, nasopharynx sa pamamagitan ng mga sensitibong sanga ng trigeminal at glossopharyngeal nerves ay pumapasok sa nuclei ng medulla oblongata, mula dito ang utos. papunta sa motor nuclei ng trigeminal, vagus, facial, glossopharyngeal, accessory o hypoglossal nerves, bilang isang resulta, ang isa o isa pang proteksiyon na reflex ay natanto. Sa parehong paraan, dahil sa sunud-sunod na pagsasama ng mga grupo ng kalamnan ng ulo, leeg, dibdib at dayapragm, ang mga reflexes ng pag-uugali sa pagkain ay nakaayos: pagsuso, pagnguya, paglunok.
Bilang karagdagan, ang medulla oblongata ay nag-aayos ng posture maintenance reflexes. Ang mga reflexes na ito ay nabuo dahil sa afferentation mula sa mga receptor sa vestibule ng cochlea at kalahating bilog na mga kanal sa superior vestibular nucleus; mula dito, ang naprosesong impormasyon upang masuri ang pangangailangan para sa pagbabago sa pustura ay ipinadala sa lateral at medial vestibular nuclei. Ang mga nuclei na ito ay kasangkot sa pagtukoy kung aling mga sistema ng kalamnan, mga segment ng spinal cord ang dapat makibahagi sa pagbabago ng pustura, samakatuwid, mula sa mga neuron ng medial at lateral nuclei sa kahabaan ng vestibulospinal pathway, ang signal ay papunta sa mga anterior horn ng kaukulang mga segment. ng spinal cord, innervating ang mga kalamnan, na ang pakikilahok sa pagbabago ng pustura sa sandaling ito ay kinakailangan.
Ang pagbabago ng postura ay isinasagawa dahil sa static at statokinetic reflexes. Ang mga static na reflexes ay kumokontrol sa tono ng kalamnan ng kalansay upang mapanatili ang isang tiyak na posisyon ng katawan. Ang mga statokinetic reflexes ng medulla oblongata ay nagbibigay ng muling pamamahagi ng tono ng mga kalamnan ng trunk upang ayusin ang isang pose na naaayon sa sandali ng rectilinear o rotational na paggalaw.
Karamihan sa mga autonomic reflexes ng medulla oblongata ay natanto sa pamamagitan ng nuclei ng vagus nerve na matatagpuan dito, na tumatanggap ng impormasyon tungkol sa estado ng aktibidad ng puso, mga daluyan ng dugo, digestive tract, baga, digestive glands, atbp. Bilang tugon sa ang impormasyong ito, ang nuclei ay nag-aayos ng mga reaksyon ng motor at secretory ng mga organ na ito.
Ang paggulo ng nuclei ng vagus nerve ay nagdudulot ng pagtaas sa pag-urong ng makinis na mga kalamnan ng tiyan, bituka, gallbladder at, sa parehong oras, pagpapahinga ng mga sphincters ng mga organo na ito. Kasabay nito, ang gawain ng puso ay bumagal at humina, ang lumen ng bronchi ay makitid.
Ang aktibidad ng nuclei ng vagus nerve ay ipinahayag din sa isang pagtaas sa pagtatago ng bronchial, gastric, bituka glandula, sa paggulo ng pancreas, secretory cells ng atay.
Sa medulla oblongata, ang sentro ng paglalaway ay naisalokal, ang parasympathetic na bahagi nito ay nagbibigay ng pagtaas sa kabuuang pagtatago, at ang nagkakasundo na bahagi ng pagtatago ng protina ng mga glandula ng salivary.
Ang mga sentro ng paghinga at vasomotor ay matatagpuan sa istraktura ng reticular formation ng medulla oblongata. Ang kakaiba ng mga sentrong ito ay ang kanilang mga neuron ay nasasabik nang reflexively at sa ilalim ng impluwensya ng chemical stimuli.
Ang sentro ng paghinga ay naisalokal sa medial na bahagi ng reticular formation ng bawat simetriko kalahati ng medulla oblongata at nahahati sa dalawang bahagi, paglanghap at pagbuga.
Sa reticular formation ng medulla oblongata, isa pang mahalagang sentro ang kinakatawan - ang vasomotor center (regulasyon ng vascular tone). Gumagana ito kasabay ng mga nakapatong na istruktura ng utak at, higit sa lahat, sa hypothalamus. Ang paggulo ng sentro ng vasomotor ay palaging nagbabago sa ritmo ng paghinga, tono ng bronchi, mga kalamnan ng bituka, pantog ng ihi, kalamnan ng ciliary, atbp. Ito ay dahil sa katotohanan na ang reticular formation ng medulla oblongata ay may synaptic na koneksyon sa hypothalamus at iba pang mga sentro.
Sa gitnang mga seksyon ng reticular formation, may mga neuron na bumubuo sa reticulospinal pathway, na may nagbabawal na epekto sa mga motor neuron ng spinal cord. Sa ilalim ng IV ventricle, matatagpuan ang mga neuron ng "blue spot". Ang kanilang tagapamagitan ay norepinephrine. Isinaaktibo ng mga neuron na ito ang reticulospinal pathway sa panahon ng pagtulog ng REM, na humahantong sa pagsugpo sa mga spinal reflexes at pagbaba sa tono ng kalamnan.
Sintomas ng pinsala. Ang pinsala sa kaliwa o kanang kalahati ng medulla oblongata sa itaas ng intersection ng pataas na mga landas ng proprioceptive sensitivity ay nagdudulot ng mga kaguluhan sa sensitivity at trabaho ng mga kalamnan ng mukha at ulo sa gilid ng pinsala. Kasabay nito, sa kabaligtaran na bahagi na nauugnay sa gilid ng pinsala, may mga karamdaman ng sensitivity ng balat at paralisis ng motor ng puno ng kahoy at mga paa. Ito ay dahil sa ang katunayan na ang pataas at pababang mga landas mula sa spinal cord at papunta sa spinal cord ay nagsalubong, at ang nuclei ng cranial nerves ay nagpapapasok sa kanilang kalahati ng ulo, ibig sabihin, ang cranial nerves ay hindi nagsalubong.
tulay
Ang tulay (ponscerebri, ponsVarolii) ay matatagpuan sa itaas ng medulla oblongata at gumaganap ng sensory, conductive, motor, integrative reflex functions.
Kasama sa tulay ang nuclei ng facial, trigeminal, abducent, vestibular at cochlear nerves (vestibular at cochlear nuclei), ang nuclei ng vestibular cochlear nerve (vestibular nerve): lateral (Deiters) at superior (Bekhterev). Ang reticular formation ng pons ay malapit na nauugnay sa reticular formation ng medulla at medulla oblongata.
Ang isang mahalagang istraktura ng pons ay ang gitnang pedicle ng cerebellum. Siya ang nagbibigay ng functional compensatory at morphological na koneksyon ng cerebral cortex sa cerebellar hemispheres.
Ang mga sensory function ng pons ay ibinibigay ng nuclei ng vestibular cochlear at trigeminal nerves. Ang cochlear na bahagi ng vestibular cochlear nerve ay nagtatapos sa utak sa cochlear nuclei; ang vestibular na bahagi ng vestibular cochlear nerve - sa triangular nucleus, ang Deiters nucleus, ang ankylosing spondylitis nucleus. Dito nagaganap ang pangunahing pagsusuri ng vestibular stimuli ng kanilang lakas at direksyon.
Ang sensory nucleus ng trigeminal nerve ay tumatanggap ng mga signal mula sa mga receptor ng balat ng mukha, anterior na anit, mauhog lamad ng ilong at bibig, ngipin at conjunctiva ng eyeball. Ang facial nerve (n. Facialis) ay nagpapaloob sa lahat ng facial muscles ng mukha. Ang abducens nerve (n. Abducens) ay nagpapapasok sa rectus lateral na kalamnan, na dumudukot sa eyeball palabas.
Ang motor na bahagi ng trigeminal nerve nucleus (n. Trigeminus) ay nagpapaloob sa mga kalamnan ng nginunguyang, ang kalamnan na humihila sa eardrum, at ang kalamnan na humihila sa palatine curtain.
Conductive function ng tulay. Binibigyan ng longitudinally at transversely arranged fibers. Ang mga transversely located fibers ay bumubuo sa itaas at mas mababang mga layer, at sa pagitan ng mga ito ay may mga pyramidal pathway na umaabot mula sa cerebral cortex. Neural clusters - ang nuclei ng pons - ay matatagpuan sa pagitan ng mga transverse fibers. Mula sa kanilang mga neuron, nagsisimula ang mga transverse fibers, na papunta sa kabaligtaran ng tulay, na bumubuo sa gitnang pedicle ng cerebellum at nagtatapos sa cortex nito.
Sa takip ng tulay, may mga longitudinal na pagpapalawak ng mga bundle ng mga hibla ng medial loop (lemniscusmedialis). Ang mga ito ay tinawid ng mga transverse fibers ng trapezoidal body (corpustrapezoideum), na mga axon ng cochlear na bahagi ng vestibular cochlear nerve ng kabaligtaran na bahagi, na nagtatapos sa nucleus ng superior olive (olivasuperior). Mula sa nucleus na ito ay may mga landas ng lateral loop (lemniscus lateralis), na ipinadala sa posterior quadruple ng midbrain at sa medial geniculate na katawan ng diencephalon.
Ang anterior at posterior nuclei ng trapezoidal body at ang lateral loop ay naisalokal sa tectum. Ang mga nuclei na ito, kasama ang superior olive, ay nagbibigay ng pangunahing pagsusuri ng impormasyon mula sa organ ng pandinig at pagkatapos ay nagpapadala ng impormasyon sa posterior tubercles ng quadruples.
Ang tectum ay naglalaman din ng mahabang medial at tectospinal pathways.
Ang tamang mga neuron ng istraktura ng pons ay bumubuo sa reticular formation nito, ang nucleus ng facial, abducens nerves, ang motor na bahagi ng nucleus at ang gitnang sensory nucleus ng trigeminal nerve.
Ang reticular formation ng pons ay isang pagpapatuloy ng reticular formation ng medulla oblongata at ang simula ng parehong sistema ng midbrain. Ang mga axon ng mga neuron sa pons reticular formation ay pumupunta sa cerebellum, sa spinal cord (reticulospinal pathway). Ang huli ay nagpapagana ng mga neuron ng spinal cord.
Ang pons reticularis ay nakakaapekto sa cerebral cortex, na nagiging sanhi ng paggising o pagtulog nito. Sa reticular formation ng pons mayroong dalawang grupo ng nuclei na kabilang sa karaniwang respiratory center. Ang isang sentro ay nagpapagana sa sentro ng inspirasyon ng medulla oblongata, ang isa naman ay nagpapagana ng sentro ng pag-alis. Ang mga neuron ng respiratory center na matatagpuan sa tulay ay umaangkop sa gawain ng mga respiratory cells ng medulla oblongata alinsunod sa pagbabago ng estado ng katawan.
8) (Midbrain, mga pag-andar nito, pakikilahok sa regulasyon ng tono ng kalamnan)
Morphofunctional na organisasyon. Ang midbrain (mesencephalon) ay kinakatawan ng quadruple at ang mga binti ng utak. Ang pinakamalaking nuclei ng midbrain ay ang pulang nucleus, ang substantia nigra at ang nuclei ng cranial (oculomotor at block) nerves, pati na rin ang nucleus ng reticular formation.
Mga function ng pandama. Ang mga ito ay natanto dahil sa pagtanggap ng visual at auditory na impormasyon sa loob nito.
Conductive function. Binubuo ito sa katotohanan na ang lahat ng mga pataas na landas ay dumadaan dito sa nakapatong na thalamus (medial loop, spinothalamic pathway), ang malaking utak at ang cerebellum. Ang mga pababang landas ay dumadaan sa midbrain patungo sa medulla oblongata at spinal cord. Ito ang pyramidal path, cortical-bridge fibers, rubroreticulospinal path.
Pag-andar ng motor. Ito ay natanto dahil sa nucleus ng block nerve (n. Trochlearis), ang nuclei ng oculomotor nerve (item oculomotorius), ang red nucleus (nucleus ruber), ang black matter (substantianigra).
Ang pulang nuclei ay matatagpuan sa tuktok ng cerebral peduncles. Ang mga ito ay nauugnay sa cerebral cortex (mga landas na bumababa mula sa cortex), subcortical nuclei, cerebellum, spinal cord (red-nuclear-spinal pathway). Ang basal ganglia ng utak, ang cerebellum ay may mga dulo sa pulang nuclei. Ang pagkagambala sa mga koneksyon ng pulang nuclei na may reticular formation ng medulla oblongata ay humahantong sa decerebrational rigidity. Ang kundisyong ito ay nailalarawan sa pamamagitan ng malakas na pag-igting ng mga extensor na kalamnan ng mga paa, leeg, at likod. Ang pangunahing dahilan para sa paglitaw ng decerebrational rigidity ay ang binibigkas na activating effect ng lateral vestibular nucleus (Deiters nucleus) sa extensor motor neurons. Ang impluwensyang ito ay pinakamataas sa kawalan ng nagbabawal na mga impluwensya ng pulang nucleus at nakapatong na mga istraktura, pati na rin ang cerebellum. Kapag ang utak ay nailipat sa ibaba ng nucleus ng lateral vestibular nerve, nawawala ang decerebral rigidity.
Ang pulang nuclei, na tumatanggap ng impormasyon mula sa motor zone ng cerebral cortex, subcortical nuclei at cerebellum tungkol sa paparating na paggalaw at ang estado ng musculoskeletal system, ay nagpapadala ng mga corrective impulses sa mga motor neuron ng spinal cord kasama ang rubrospinal tract at sa gayon ay kinokontrol ang kalamnan. tono, inihahanda ang antas nito para sa nakabalangkas na boluntaryong kilusan ...
Ang isa pang functional na mahalagang nucleus ng midbrain - ang itim na substansiya - ay matatagpuan sa mga binti ng utak, kinokontrol ang mga kilos ng pagnguya, paglunok (kanilang pagkakasunud-sunod), at tinitiyak ang tumpak na paggalaw ng mga daliri ng kamay, halimbawa, kapag nagsusulat. Ang mga neuron ng nucleus na ito ay nagagawang synthesize ang mediator dopamine, na ibinibigay ng axonal transport sa basal ganglia ng utak. Ang pagkatalo ng substantia nigra ay humahantong sa isang paglabag sa plastik na tono ng mga kalamnan. Ang pinong regulasyon ng plastik na tono kapag naglalaro ng biyolin, nagsusulat, gumaganap ng mga graphic na gawa ay ibinibigay ng itim na substansiya. Kasabay nito, na may pangmatagalang pagpapanatili ng isang tiyak na pustura, ang mga pagbabago sa plastik ay nangyayari sa mga kalamnan dahil sa mga pagbabago sa kanilang mga koloidal na katangian, na nagsisiguro sa pinakamababang pagkonsumo ng enerhiya. Ang regulasyon ng prosesong ito ay isinasagawa ng mga selula ng substantia nigra.
Kinokontrol ng mga neuron ng nuclei ng oculomotor at trochlear nerve ang paggalaw ng mata pataas, pababa, palabas, papunta sa ilong at pababa sa sulok ng ilong. Ang mga neuron ng accessory nucleus ng oculomotor nerve (Yakubovich's nucleus) ay kinokontrol ang lumen ng mag-aaral at ang curvature ng lens.
Mga function ng reflex. Ang functionally independent structures ng midbrain ay ang mga hillocks ng quadruple. Ang mga nasa itaas ay ang pangunahing mga subcortical center ng visual analyzer (kasama ang mga lateral geniculate na katawan ng diencephalon), ang mga mas mababa ay ang auditory (kasama ang medial geniculate na katawan ng diencephalon). Sa kanila, ang pangunahing paglipat ng visual at auditory na impormasyon ay nagaganap. Mula sa mga tubercle ng quadruple, ang mga axon ng kanilang mga neuron ay napupunta sa reticular formation ng trunk, ang mga motor neuron ng spinal cord. Ang mga quadruple neuron ay maaaring polymodal at detector. Sa huling kaso, sila ay tumutugon sa isang senyales lamang ng pangangati, halimbawa, isang pagbabago sa liwanag at kadiliman, ang direksyon ng paggalaw ng isang pinagmumulan ng liwanag, atbp. Ang pag-activate ng midbrain sa mga kasong ito sa pamamagitan ng hypothalamus ay humahantong sa pagtaas ng tono ng kalamnan, pagtaas ng rate ng puso; mayroong paghahanda para sa pag-iwas, para sa isang nagtatanggol na reaksyon.
Ang quadruple ay nag-aayos ng orientational visual at auditory reflexes.
Sa mga tao, ang quadruple reflex ay isang sentinel reflex. Sa mga kaso ng pagtaas ng excitability ng quadruples na may biglaang tunog o light stimulation, ang isang tao ay nagsisimulang mag-flinching, kung minsan ay tumatalon sa kanilang mga paa, sumisigaw, pinakamabilis na lumalayo sa stimulus, at kung minsan ay tumakas.
Kung ang quadruple reflex ay nabalisa, ang isang tao ay hindi maaaring mabilis na lumipat mula sa isang uri ng paggalaw patungo sa isa pa. Dahil dito, ang quadruples ay nakikibahagi sa organisasyon ng mga boluntaryong kilusan.
Brainstem reticular formation
Ang reticular formation (formatioreticularis; RF) ng utak ay kinakatawan ng isang network ng mga neuron na may maraming nagkakalat na koneksyon sa pagitan nila at sa halos lahat ng mga istruktura ng central nervous system. Ang RF ay matatagpuan sa kapal ng gray matter ng medulla oblongata, midbrain, at diencephalon at sa una ay nauugnay sa RF ng spinal cord. Sa bagay na ito, ipinapayong isaalang-alang ito bilang pinag-isang sistema... Ang mga koneksyon sa network ng mga neuron ng RF sa kanilang mga sarili ay nagpapahintulot sa Deiters na tawagan itong reticular formation ng utak.
Ang RF ay may direktang at feedback na mga koneksyon sa cerebral cortex, basal ganglia, diencephalon, cerebellum, gitna, medulla oblongata at spinal cord.
Ang pangunahing tungkulin ng RF ay upang ayusin ang antas ng aktibidad ng cerebral cortex, cerebellum, thalamus, at spinal cord.
Sa isang banda, ang pangkalahatang katangian ng impluwensya ng RF sa maraming istruktura ng utak ay nagbigay ng batayan upang ituring itong isang hindi tiyak na sistema. Gayunpaman, ang mga pag-aaral na may pagpapasigla ng RF trunk ay nagpakita na maaari itong piliing magsagawa ng activating o inhibitory effect sa iba't ibang anyo ng pag-uugali, sa sensory, motor, at visceral system ng utak. Tinitiyak ng istraktura ng network ang mataas na pagiging maaasahan ng paggana ng RF, paglaban sa mga nakakapinsalang impluwensya, dahil ang lokal na pinsala ay palaging binabayaran ng natitirang mga elemento ng network. Sa kabilang banda, ang mataas na pagiging maaasahan ng paggana ng RF ay sinisiguro ng katotohanan na ang pangangati ng alinman sa mga bahagi nito ay makikita sa aktibidad ng buong RF ng isang naibigay na istraktura dahil sa diffuseness ng mga koneksyon.
Karamihan sa mga RF neuron ay may mahabang dendrite at isang maikling axon. May mga higanteng neuron na may mahabang axon na bumubuo ng mga landas mula sa RF patungo sa iba pang bahagi ng utak, halimbawa, sa ibaba ng agos, reticulospinal, at rubrospinal. Ang mga axon ng RF neuron ay bumubuo ng isang malaking bilang ng mga collateral at synapses, na nagtatapos sa mga neuron ng iba't ibang bahagi ng utak. Ang mga axon ng RF neuron, papunta sa cerebral cortex, ay nagtatapos dito sa mga dendrite ng mga layer I at II.
Ang aktibidad ng mga neuron ng RF ay naiiba at, sa prinsipyo, ay katulad ng aktibidad ng mga neuron sa iba pang mga istruktura ng utak, ngunit kabilang sa mga neuron ng RF mayroong mga may isang matatag na aktibidad ng ritmo na hindi nakasalalay sa mga papasok na signal.
Kasabay nito, sa RF ng midbrain at pons mayroong mga neuron na "tahimik" sa pamamahinga, iyon ay, hindi sila bumubuo ng mga impulses, ngunit nasasabik kapag ang mga visual o auditory receptor ay pinasigla. Ito ang tinatawag na mga partikular na neuron na nagbibigay ng mabilis na pagtugon sa biglaang, hindi nakikilalang mga signal. Ang isang makabuluhang bilang ng mga RF neuron ay polysensory.
Sa RF ng medulla oblongata, ang midbrain at ang pons, ang mga signal ng iba't ibang mga sensasyon ay nagtatagpo. Ang mga neuron ng tulay ay tumatanggap ng mga signal pangunahin mula sa mga sistema ng somatosensory. Ang mga signal mula sa visual at auditory sensory system ay pangunahing dumarating sa mga RF neuron ng midbrain.
Kinokontrol ng RF ang paghahatid ng pandama na impormasyon na dumadaan sa nuclei ng thalamus, dahil sa ang katunayan na sa matinding panlabas na pagpapasigla, ang mga neuron ng hindi tiyak na nuclei ng thalamus ay pinipigilan, sa gayon ay inaalis ang kanilang epekto sa pagbabawal mula sa relay nuclei ng parehong thalamus at pinapadali ang paghahatid ng pandama na impormasyon sa cerebral cortex.
Sa RF ng pons, medulla oblongata, may mga neuron na tumutugon sa masakit na stimuli mula sa mga kalamnan o panloob na organo, na lumilikha ng isang pangkalahatang nagkakalat na hindi komportable, hindi palaging malinaw na naisalokal, masakit na sensasyon ng "mapurol na sakit".
Ang pag-uulit ng anumang uri ng pagpapasigla ay humahantong sa pagbawas sa aktibidad ng salpok ng mga RF neuron, ibig sabihin, ang mga proseso ng adaptasyon (addiction) ay likas sa mga RF neuron ng stem ng utak.
Ang RF ng stem ng utak ay direktang nauugnay sa regulasyon ng tono ng kalamnan, dahil ang RF ng stem ng utak ay tumatanggap ng mga signal mula sa mga visual at vestibular analyzer at ang cerebellum. Ang mga signal na nag-aayos ng posisyon ng ulo, puno ng kahoy, atbp., ay ipinapadala mula sa RF sa mga motor neuron ng spinal cord at ang nuclei ng cranial nerves.
Ang mga reticular pathway na nagpapadali sa aktibidad ng mga sistema ng motor ng spinal cord ay nagmumula sa lahat ng bahagi ng RF. Ang mga pathway na humahantong mula sa tulay ay humahadlang sa aktibidad ng spinal cord motor neuron na nagpapasigla sa mga flexor na kalamnan at nagpapagana sa mga motor neuron ng mga extensor na kalamnan. Ang mga landas na humahantong mula sa RF ng medulla oblongata ay may kabaligtaran na mga epekto. Ang pangangati ng RF ay humahantong sa panginginig, pagtaas ng tono ng kalamnan. Matapos ang pagwawakas ng pangangati, ang epekto na dulot nito ay nagpapatuloy sa loob ng mahabang panahon, tila dahil sa sirkulasyon ng paggulo sa network ng mga neuron.
Ang brain stem RF ay kasangkot sa paghahatid ng impormasyon mula sa cerebral cortex, spinal cord sa cerebellum at, sa kabaligtaran, mula sa cerebellum patungo sa parehong mga sistema. Ang pag-andar ng mga koneksyon na ito ay ang paghahanda at pagpapatupad ng mga kasanayan sa motor na nauugnay sa habituation, mga reaksyon sa pag-orient, mga reaksyon ng sakit, samahan ng paglalakad, at paggalaw ng mata.
Ang regulasyon ng autonomic na aktibidad ng RF organism ay inilarawan sa Seksyon 4.3, ngunit dito napapansin namin na ang regulasyong ito ay pinaka-malinaw na ipinahayag sa paggana ng respiratory at cardiovascular centers. Sa regulasyon ng mga autonomic function, ang tinatawag na panimulang mga neuron ng RF ay may malaking kahalagahan. Nagbibigay sila ng sirkulasyon ng paggulo sa loob ng isang pangkat ng mga neuron, na nagbibigay ng tono ng mga regulated autonomic system.
Ang mga impluwensya ng RF ay maaaring malawak na nahahati sa pababa at pataas. Sa turn, ang bawat isa sa mga impluwensyang ito ay may nagbabawal at nakapagpapasigla na epekto.
Ang mga pataas na epekto ng RF sa cerebral cortex ay nagpapataas ng tono nito, kinokontrol ang excitability ng mga neuron nito, nang hindi binabago ang pagtitiyak ng mga tugon sa sapat na stimuli. Naiimpluwensyahan ng RF ang functional state ng lahat ng sensory regions ng utak, samakatuwid, ito ay gumaganap ng isang papel sa pagsasama ng sensory information mula sa iba't ibang analyzer.
Direktang nauugnay ang RF sa regulasyon ng cycle ng wakefulness-sleep. Ang pagpapasigla ng ilang istruktura ng RF ay humahantong sa pag-unlad ng pagtulog, habang ang pagpapasigla ng iba ay nagdudulot ng paggising. Iniharap nina G. Magun at D. Moruzzi ang konsepto ayon sa kung saan ang lahat ng uri ng signal na nagmumula sa mga peripheral receptor ay umaabot sa medulla oblongata at sa pons sa pamamagitan ng RF collaterals, kung saan lumipat sila sa mga neuron na nagbibigay ng mga pataas na daanan patungo sa thalamus at pagkatapos ay sa cerebral cortex.
Ang paggulo ng RF ng medulla oblongata o pons ay nagiging sanhi ng pag-synchronize ng aktibidad ng cerebral cortex, ang hitsura ng mabagal na ritmo sa mga de-koryenteng parameter nito, at pagsugpo sa pagtulog.
Ang paggulo ng RF ng midbrain ay nagiging sanhi ng kabaligtaran na epekto ng paggising: desynchronization ng electrical activity ng cortex, ang hitsura ng mabilis na low-amplitude β-like rhythms sa electroencephalogram.
Ipinakita ni G. Bremer (1935) na kung ang utak ay pinutol sa pagitan ng anterior at posterior tubercles ng quadruple, ang hayop ay hihinto sa pagtugon sa lahat ng uri ng signal; kung ang hiwa ay ginawa sa pagitan ng medulla oblongata at ng midbrain (habang ang RF ay nagpapanatili ng koneksyon sa forebrain), pagkatapos ay ang hayop ay tumutugon sa liwanag, tunog, at iba pang mga signal. Dahil dito, ang pagpapanatili ng isang aktibong pagsusuri sa estado ng utak ay posible habang pinapanatili ang koneksyon sa forebrain.
Ang reaksyon ng pag-activate ng cerebral cortex ay sinusunod sa pagpapasigla ng RF ng medulla oblongata, midbrain, at diencephalon. Kasabay nito, ang pangangati ng ilang nuclei ng thalamus ay humahantong sa paglitaw ng mga limitadong lokal na lugar ng paggulo, at hindi sa pangkalahatang paggulo nito, tulad ng kaso sa pangangati ng ibang bahagi ng RF.
Ang brain stem RF ay maaaring magkaroon ng hindi lamang isang kapana-panabik, kundi pati na rin ang isang nagbabawal na epekto sa aktibidad ng cerebral cortex.
Ang mga pababang epekto ng RF ng stem ng utak sa aktibidad ng regulasyon ng spinal cord ay itinatag ni I.M.Sechenov (1862). Ipinakita niya na kapag ang midbrain ay pinasigla ng mga kristal ng asin sa isang palaka, ang mga paw withdrawal reflexes ay lumilitaw nang dahan-dahan, nangangailangan ng mas matinding pagpapasigla, o hindi lumilitaw sa lahat, iyon ay, sila ay inhibited.
Nalaman ni G. Magun (1945-1950), na nagdudulot ng mga lokal na iritasyon sa RF ng medulla oblongata, na kapag ang ilang mga punto ay pinasigla, ang mga flexion reflexes ng front paw, tuhod, at corneal reflexes ay nagiging inhibited at nagiging tamad. Sa pagpapasigla ng RF sa iba pang mga punto ng medulla oblongata, ang mga parehong reflexes na ito ay mas madaling mapukaw, ay mas malakas, ibig sabihin, ang kanilang pagpapatupad ay pinadali. Ayon kay Megun, tanging ang RF ng medulla oblongata ang makakapagbigay ng mga epektong nagbabawal sa mga reflexes ng spinal cord, at ang mga impluwensyang nagpapadali ay kinokontrol ng buong RF ng trunk at spinal cord.
9) (Cerebellum, ang pakikilahok nito sa regulasyon ng mga pag-andar ng motor at autonomic)
Ang cerebellum (cerebellum, maliit na utak) ay isa sa mga pinagsama-samang istruktura ng utak, na nakikibahagi sa koordinasyon at regulasyon ng mga boluntaryo, hindi sinasadyang paggalaw, sa regulasyon ng mga autonomic at pag-uugali na pag-andar.
Mga tampok ng morphofunctional na organisasyon at komunikasyon ng cerebellum. Ang pagpapatupad ng mga function na ito ay ibinibigay ng mga sumusunod na morphological features ng cerebellum:
1) ang cerebellar cortex ay binuo nang pantay-pantay, may mga stereotyped na koneksyon, na lumilikha ng mga kondisyon para sa mabilis na pagproseso ng impormasyon;
2) ang pangunahing elemento ng neural ng cortex - ang Purkinje cell, ay may malaking bilang ng mga input at bumubuo ng isang solong axonal output mula sa cerebellum, ang mga collateral na nagtatapos sa mga istrukturang nuklear nito;
3) halos lahat ng uri ng sensory stimuli ay ipinapalabas sa mga selula ng Purkinje: proprioceptive, balat, visual, auditory, vestibular, atbp.;
4) ang mga output mula sa cerebellum ay nagbibigay ng mga koneksyon nito sa cerebral cortex, kasama ang mga stem formation at ang spinal cord.
Ang cerebellum ay anatomically at functionally nahahati sa luma, sinaunang at bagong mga bahagi.
Ang lumang bahagi ng cerebellum (archicerebellum) - ang vestibular cerebellum - ay kinabibilangan ng clumpy-floccular lobe. Ang bahaging ito ay may pinaka binibigkas na mga koneksyon sa vestibular analyzer, na nagpapaliwanag ng kahalagahan ng cerebellum sa regulasyon ng balanse.
Ang sinaunang bahagi ng cerebellum (paleocerebellum) - ang spinal cerebellum - ay binubuo ng mga seksyon ng vermis at ang pyramid ng cerebellum, uvula, pericellular region at tumatanggap ng impormasyon higit sa lahat mula sa proprioceptive system ng mga kalamnan, tendon, periosteum, joint membranes.
Ang bagong cerebellum (neocerebellum) ay kinabibilangan ng cortex ng cerebellar hemispheres at mga lugar ng worm; tumatanggap siya ng impormasyon mula sa cortex, pangunahin sa kahabaan ng frontal-cerebellopontine pathway, mula sa visual at auditory receptive system, na nagpapahiwatig ng kanyang pakikilahok sa pagsusuri ng visual at auditory signal at ang organisasyon ng mga reaksyon sa kanila.
Ang cerebellar cortex ay may isang tiyak na istraktura na hindi nauulit kahit saan sa central nervous system. Ang upper (I) layer ng cerebellar cortex - ang molecular layer, ay binubuo ng mga parallel fibers, dendritic ramifications at axons ng mga layer II at III. Sa ibabang bahagi ng molecular layer, matatagpuan ang mga basket at stellate cell, na tinitiyak ang pakikipag-ugnayan ng mga selula ng Purkinje.
Ang gitnang (II) layer ng cortex ay nabuo ng mga Purkinje cells, na nakaayos sa isang hilera at may pinakamakapangyarihang dendritic system sa central nervous system. Sa dendritic field ng isang Purkinje cell, maaaring magkaroon ng hanggang 60,000 synapses. Dahil dito, isinasagawa ng mga cell na ito ang gawain ng pagkolekta, pagproseso at pagpapadala ng impormasyon. Ang mga axon ng mga selula ng Purkinje ay ang tanging paraan kung saan ang cerebellar cortex ay nagpapadala ng impormasyon sa nuclei nito at sa nucleus ng istraktura ng cerebellar.
Sa ilalim ng II layer ng cortex (sa ilalim ng mga selula ng Purkinje) mayroong isang butil-butil (III) na layer, na binubuo ng mga butil na selula, ang bilang nito ay umabot sa 10 bilyon. Ang mga axon ng mga selulang ito ay tumaas paitaas, nahahati sa isang T-hugis sa ang ibabaw ng cortex, na bumubuo ng mga landas ng mga contact na may mga selula ng Purkinje. Narito ang mga selulang Golgi.
Mula sa cerebellum, ang impormasyon ay umaalis sa itaas at ibabang mga binti. Sa pamamagitan ng itaas na mga binti, ang mga signal ay napupunta sa thalamus, ang pons, ang pulang nucleus, ang nuclei ng stem ng utak, at ang reticular formation ng midbrain. Sa pamamagitan ng mas mababang mga binti ng cerebellum, ang mga senyales ay napupunta sa medulla oblongata sa kanyang vestibular nuclei, olives, at reticular formation. Ang gitnang pedicles ng cerebellum ay nagkokonekta sa bagong cerebellum sa frontal lobe ng utak.
Ang aktibidad ng impulse ng mga neuron ay naitala sa layer ng Purkinje cells at granular layer, at ang dalas ng pagbuo ng mga impulses ng mga cell na ito ay mula 20 hanggang 200 bawat segundo. Ang mga cell ng cerebellar nuclei ay gumagawa ng mga impulses na mas madalas - 1-3 impulses bawat segundo.
Ang pagpapasigla ng itaas na layer ng cerebellar cortex ay humahantong sa pangmatagalang (hanggang 200 ms) na pagsugpo sa aktibidad ng cell ng Purkinje. Ang parehong pagsugpo ay nangyayari sa mga signal ng liwanag at tunog. Ang kabuuang mga pagbabago sa aktibidad ng elektrikal ng cerebellar cortex hanggang sa pangangati ng sensory nerve ng anumang kalamnan ay lumilitaw sa anyo ng isang positibong oscillation (pagbawal sa aktibidad ng cortex, hyperpolarization ng Purkinje cells), na nangyayari pagkatapos ng 15-20 ms at tumatagal ng 20-30 ms, pagkatapos nito ay nangyayari ang isang excitation wave, na tumatagal ng hanggang 500 ms (depolarization ng Purkinje cells).
Sa cerebellar cortex mula sa mga receptor ng balat, kalamnan, articular membrane, periosteum, ang mga signal ay pumapasok sa tinatawag na spinal-cerebellar tracts: kasama ang posterior (dorsal) at anterior (ventral). Ang mga landas na ito patungo sa cerebellum ay dumadaan sa mababang olive ng medulla oblongata. Mula sa mga olive cell, mayroong tinatawag na climbing fibers, na sumasanga sa mga dendrite ng Purkinje cells.
Ang nuclei ng pons ay nagpapadala ng mga afferent pathway sa cerebellum, na bumubuo ng mga mossy fibers na nagtatapos sa mga butil na selula ng layer III ng cerebellar cortex. Mayroong koneksyon sa pagitan ng cerebellum at ang mala-bughaw na lugar ng midbrain gamit ang mga adrenergic fibers. Ang mga fibers na ito ay nakakapaglabas ng norepinephrine sa intercellular space ng cerebellar cortex, at sa gayon ay nakakatawang binabago ang estado ng excitability ng mga selula nito.
Ang mga axon ng mga selula sa layer III ng cerebellar cortex ay nagdudulot ng pagsugpo sa mga selula ng Purkinje at mga selula ng butil ng kanilang sariling layer.
Ang mga purkinje cell, naman, ay pumipigil sa aktibidad ng mga neuron sa cerebellar nuclei. Ang cerebellar nuclei ay may mataas na tonic na aktibidad at kinokontrol ang tono ng isang bilang ng mga sentro ng motor sa intermediate, middle, oblong, at spinal cord.
Ang subcortical system ng cerebellum ay binubuo ng tatlong functionally different nuclear formations: ang nucleus ng tent, corky, spherical at dentate nuclei.
Ang tent nucleus ay tumatanggap ng impormasyon mula sa medial zone ng cerebellar cortex at konektado sa Deiters nucleus at ang RF ng medulla oblongata at midbrain. Mula dito, ang mga signal ay naglalakbay kasama ang reticulospinal pathway patungo sa mga motor neuron ng spinal cord.
Ang intermediate cerebellar cortex ay inaasahang papunta sa corky at globular nuclei. Mula sa kanila, ang mga koneksyon ay pumupunta sa midbrain sa pulang nucleus, pagkatapos ay sa spinal cord kasama ang rubrospinal pathway. Ang pangalawang landas mula sa intermediate nucleus ay papunta sa thalamus at higit pa sa motor area ng cerebral cortex.
Ang dentate nucleus, na tumatanggap ng impormasyon mula sa lateral zone ng cerebellar cortex, ay konektado sa thalamus, at sa pamamagitan nito - kasama ang motor zone ng cerebellar cortex.
Kontrol ng cerebellar ng aktibidad ng motor. Ang mga efferent signal mula sa cerebellum hanggang sa spinal cord ay kumokontrol sa lakas ng mga contraction ng kalamnan, nagbibigay ng kakayahan para sa matagal na tonic na pag-urong ng kalamnan, ang kakayahang mapanatili ang pinakamainam na tono ng kalamnan sa pahinga o sa panahon ng paggalaw, upang masukat ang mga boluntaryong paggalaw para sa layunin ng paggalaw na ito, upang mabilis na lumipat mula sa pagbaluktot patungo sa extension at vice versa.
Ang cerebellum ay nagbibigay ng synergy ng mga contraction ng iba't ibang mga kalamnan sa panahon ng mga kumplikadong paggalaw. Halimbawa, ang paggawa ng isang hakbang habang naglalakad, ang isang tao ay dinadala ang kanyang binti pasulong, habang ang sentro ng grabidad ng katawan ay inilipat pasulong kasama ang paglahok ng mga kalamnan sa likod. Sa mga kasong iyon kapag ang cerebellum ay hindi natutupad ang pag-andar ng regulasyon nito, ang isang tao ay nakakaranas ng mga karamdaman ng mga pag-andar ng motor, na ipinahayag ng mga sumusunod na sintomas.
1) asthenia (astenia - kahinaan) - isang pagbawas sa lakas ng pag-urong ng kalamnan, mabilis na pagkapagod ng kalamnan;
2) astasia (astasia, mula sa Griyego a - hindi, stasia - nakatayo) - ang pagkawala ng kakayahan para sa matagal na pag-urong ng kalamnan, na nagpapahirap sa pagtayo, pag-upo, atbp.;
3) dystonia (distonia - paglabag sa tono) - isang hindi sinasadyang pagtaas o pagbaba sa tono ng kalamnan;
4) panginginig (panginginig - panginginig) - panginginig ng mga daliri, kamay, ulo sa pamamahinga; ang panginginig na ito ay tumataas sa paggalaw;
5) dysmetria (dismetria - paglabag sa panukala) - isang disorder ng pagkakapareho ng mga paggalaw, na ipinahayag alinman sa labis o hindi sapat na paggalaw. Sinusubukan ng pasyente na kumuha ng isang bagay mula sa mesa at dinadala ang kanyang kamay sa likod ng bagay (hypermetria) o hindi dinadala ito sa bagay (hypoometria);
6) ataxia (ataksia, mula sa Greek a - negation, taksia - order) - may kapansanan sa koordinasyon ng mga paggalaw. Dito, ang imposibilidad ng pagsasagawa ng mga paggalaw sa tamang pagkakasunud-sunod, sa isang tiyak na pagkakasunud-sunod, ay pinaka-malinaw na ipinahayag. Ang mga pagpapakita ng ataxia ay adiadochokinesis din, asynergia, lasing-wobbly gait. Sa adiadochokinesis, ang isang tao ay hindi mabilis na paikutin ang mga palad pataas at pababa. Sa asynergy ng mga kalamnan, hindi siya makakaupo mula sa isang nakadapa na posisyon nang walang tulong ng kanyang mga kamay. Ang lasing na lakad ay nailalarawan sa pamamagitan ng taong naglalakad na nakabuka ang mga binti, na pasuray-suray mula sa gilid patungo sa gilid mula sa linya ng paglalakad. Walang napakaraming congenital motor acts sa isang tao (halimbawa, pagsuso), ngunit natutunan niya ang karamihan sa mga paggalaw sa panahon ng kanyang buhay at nagiging awtomatiko ang mga ito.
Phylogenetically napaka sinaunang neural structure at well-developed na bahagi ng reptile brain stem. Sa una, ito ay isang mabagal na pagpapadaloy ng polysynaptic na landas na malapit na nauugnay sa mga rehiyon ng olpaktoryo at limbic. Ang progresibong pangingibabaw ng paningin at pandinig sa pandama ng pang-amoy ay humantong sa pagbabago sa sensory at motor function sa tectum ng midbrain. Ang tuwid na dorsal at teginal-spinal na mga landas ay lumalampas sa reticular formation, na pangunahing responsable para sa autonomic na regulasyon. Sa mga mammal, ang tectum, naman, ay nagsimulang gumanap ng pangalawang papel sa paghahatid ng paggulo kasama ang napakabilis na pagsasagawa ng mga hibla na nagkokonekta sa cerebral cortex na may peripheral motor at sensory neuron.
Sa utak ng tao, ang reticular formation ay nagpapanatili ng koneksyon nito sa limbic system at patuloy na gumaganap ng isang mahalagang papel sa autonomic at reflex na regulasyon.
Termino pagbuo ng reticular sumangguni lamang sa polysynaptic neural network ng brainstem, sa kabila ng katotohanan na ang network ay umaabot sa anteriorly sa thalamus at hypothalamus at posteriorly sa propriospinal tract ng spinal cord.
Pangkalahatang istraktura ipinapakita sa figure sa ibaba. Ang median reticular formation ay nabuo sa pamamagitan ng isang bilang ng suture nuclei (Greek - nuclei raphe). Karamihan sa mga serotonergic pathway ng axial na bahagi ng nervous system ay nagsisimula mula sa nuclei ng tahi.
Reticular formation (RF).(A) Mga Kagawaran. (B) Mga grupo ng aminergic at cholinergic na mga cell.
Matatagpuan sa malapit ang paramedian reticular formation. Ang seksyong ito ay ganap na binubuo ng malalaking cell neuron; sa ibabang bahagi ng pons at sa itaas na bahagi ng medulla oblongata (hanggang sa antas ng pagsasanib ng reticular formation na may gitnang reticular nucleus ng medulla oblongata), matatagpuan din ang mga higanteng cell neuron.
Ang pinakanauuna na seksyon ay isinasaalang-alang lateral small cell reticular formation... Ang mga mahahabang dendrite ng maliliit na cell neuron ay nagsasanga nang regular. Ang mga dendrite ay may nakararami na nakahalang direksyon, at sa pamamagitan ng mga puwang sa pagitan ng mga ito ay may mahahabang daanan patungo sa thalamus. Ang lateral na seksyon ay nabuo pangunahin sa pamamagitan ng mga afferent neuron. Ang mga hibla mula sa lahat ng mga sensitibong landas, kabilang ang mga pandama, ay angkop para sa kanila.
Ang mga olfactory fibers ay dumadaan sa medial forebrain bundle na matatagpuan sa tabi ng hypothalamus.
Ang mga visual na landas ay dumadaan sa itaas na punso.
Ang pandinig na mga hibla ay lumalabas mula sa itaas na core ng olibo.
Ang mga vestibular fibers ay lumalapit mula sa medial vestibular nucleus.
Ang mga somatic sensory fibers ay dumadaan sa spinal reticular tracts mula sa spinal at sariling (pangunahin o pangunahing tulay) nuclei ng trigeminal nerve.
Karamihan sa mga axon ng maliliit na selulang neuron ay masinsinang nagsasawang sa pagitan ng mga dendrite ng mga neuron ng paramedian reticular formation. Gayunpaman, ang ilan sa kanila ay bumubuo ng mga synapses na may nuclei ng cranial nerves at kasangkot sa paglikha ng mga programa sa paggalaw.
Paramedian reticular formation- nakararami ang efferent system. Ang mga axon ay medyo mahaba, ang ilan ay tumataas paitaas, na bumubuo ng mga synapses na may reticular formation ng brainstem o thalamus. Mula sa iba, ang parehong pataas at pababang mga sanga ay umaalis, na bumubuo ng isang polysynaptic na network. Ang mga hibla mula sa premotor cortex, na nagdudulot ng mga reticulospinal pathway ng pons at medulla oblongata, ay angkop para sa malalaking cell neuron.

a) Brainstem aminergic neurons... Ang mga pangkat ng aminergic (o monoaminergic) na mga neuron na nakakalat sa kahabaan ng reticular formation ay mga neuron na ang mga tagapamagitan ay nabuo mula sa mga aromatic amino acid at may ilang mga impluwensya sa cell. Ang isang grupo ay gumagawa ng neurotransmitter serotonin, tatlong iba pa ay gumagawa ng catecholamines (dopamine, norepinephrine, at adrenaline), at isang grupo ang gumagawa ng histamine.
 Mga serotonergic pathway mula sa median brainstem (suture).
Mga serotonergic pathway mula sa median brainstem (suture).
Mga neuron ng serotonin- ang pinakakaraniwang mga neuron sa anumang bahagi ng central nervous system (CNS). Kabilang dito ang mga neuron ng midbrain, ang mga hibla nito ay tumaas sa cerebral hemispheres; pontine neurons na sumasanga sa brainstem at cerebellum; mga selula ng medulla oblongata, na bumababa sa spinal cord.
Ang lahat ng bahagi ng gray matter ng central nervous system ay natatakpan ng serotonin-secreting axonal branches. Ang pagtaas sa aktibidad ng serotonergic ay ginagamit sa klinikal na kasanayan upang gamutin ang karaniwang sakit bilang pangunahing depressive disorder.

Dopaminergic neurons ng midbrain ay kinakatawan ng dalawang pangkat. May itim na substance sa junction ng gulong na may mga binti. Ang medial dito ay ang ventral tegmental nuclei, kung saan ang mesocortical fibers ay umaabot sa frontal lobe at ang mesolimbic fibers na direktang pumupunta sa nucleus accumbens.
Mga neuron na noradrenergic (norepinephrinergic). bahagyang mas kaunti kaysa sa serotonergic. Humigit-kumulang 90% ng mga katawan ng mga neuron ay puro sa locus ceruleus sa ilalim ng IV ventricle sa itaas na dulo ng pons. Nagsisimula ang mga landas mula sa asul na lugar sa lahat ng direksyon, tulad ng ipinapakita sa figure sa ibaba.
 Noradrenergic pathways mula sa pons at medulla oblongata.
Noradrenergic pathways mula sa pons at medulla oblongata.
Mga neuron na nagse-secret ng adrenaline (epinephrine-secreting). medyo kakaunti ang bilang at pangunahing matatagpuan sa mga rehiyon ng rostral / caudal ng medulla oblongata. Ang isang bahagi ng mga hibla ay umakyat sa hypothalamus, ang isa ay bumababa, na bumubuo ng mga synapses kasama ang mga preganglionic sympathetic neuron ng spinal cord.
Sa cerebral hemispheres, ang ionic at electrical activity ng aminergic neurons ay makabuluhang naiiba. Una, mayroong higit sa isang uri ng postsynaptic receptor para sa bawat amine. Pangalawa, naglalabas din ang ilang aminergic neuron mga sangkap ng protina, na may kakayahang i-regulate ang pagkilos ng tagapamagitan - bilang isang panuntunan, pagtaas ng tagal nito. Pangatlo, ang mas malalaking cortical neuron ay tumatanggap ng maraming excitatory at inhibitory na impluwensya mula sa mga lokal na nagpapalipat-lipat na excitatory network, at mayroon ding maraming iba't ibang uri ng mga receptor. Ang pag-activate ng isang uri ng aminergic receptor ay maaaring humantong sa isang malakas o mahinang epekto depende sa paunang nasasabik na estado ng neuron.
Ang aming kaalaman sa physiology at pharmacodynamics ng aminergic neurons ay malayo sa kumpleto, ngunit ang kanilang kahalagahan sa isang malawak na iba't ibang mga pag-andar ng pag-uugali ay walang pag-aalinlangan.
Bahagi ng isang cross-section sa itaas na seksyon ng tulay, na nagpapakita ng mga elemento ng reticular formation.
Ang kumplikadong istraktura ng utak ng tao ay nagpapakita ng mga lihim ng ating pag-uugali, nagpapaliwanag ng mga batas ng aktibidad ng kaisipan, ang daloy ng mga emosyon at damdamin. Ang bawat hemisphere ng utak ay may pananagutan para sa mga tiyak na pag-andar at gawain nito (halimbawa, alam na ang kanan ay responsable para sa lohika, at ang kaliwa ay para sa imahinasyon at pantasya), ngunit mayroon ding mga istruktura na nagbibigay ng isang solong at coordinated. gawain ng buong central nervous system. Ang isa sa mga istrukturang ito ay ang reticular formation.
Pangkalahatang Impormasyon
Ang reticular formation ay isang seksyon ng brainstem, na kinakatawan ng isang malawak na network ng mga nerve cell at nuclei na nag-uugnay sa iba't ibang bahagi ng utak. Hindi tulad ng iba pang mga istraktura - halimbawa, ang thalamus, hypothalamus, cerebellum - na may isang tiyak na integral na hugis (nuclei, glands), ang reticular formation ay hindi kinakatawan ng isang solong morphological formation, ngunit ito ay isang "network" (mula sa Latin reticulum - network) ng mga dendrite at axon , na, na may iba't ibang antas ng density, ay tumagos sa pagitan ng mga departamento at istruktura ng utak, na pinag-iisa ang mga ito sa isa't isa at tinitiyak ang kanilang magkasanib na aktibidad.
Metaphorically speaking: kung ang ating utak ay ipinakita sa anyo ng ilang uri ng produkto, sabihin nating, isang kamiseta, kung gayon ang reticular formation ay ang mga thread kung saan ang shirt ay natahi. Ang reticular formation ay tumagos sa mga istruktura ng medulla oblongata, midbrain at pons, ay may direktang koneksyon sa cerebellum, spinal cord, thalamus at hindi direkta sa mga nakapatong na seksyon: ang hypothalamus, visual nuclei at cortex.
Paano ito gumagana
Kasama sa reticular formation ang isang malaking bilang ng mga neuron na may branched dendrites at mahabang axon, dahil sa kung saan posible na magpadala ng mga nerve impulses sa iba't ibang bahagi ng utak at spinal cord. Sa kasong ito, maaaring makilala ang dalawang pinakamalaking grupo ng mga neural cluster:
- Ang reticulotegmental nucleus, ang mga neuron na kung saan ay tumatanggap ng mga signal mula sa mga nakapatong na bahagi ng GM (quadruple, thalamus) at ipinapadala ang mga ito sa mga istruktura ng cerebellar, at sa gayon ay kinokontrol ang ilang mahahalagang pag-andar ng motor: koordinasyon ng titig, paggalaw ng mata.
- Ang lateral nucleus, na ang mga neuron ay umakyat mula sa mga istruktura ng spinal cord at vestibular nuclei at nagbibigay ng impormasyon sa GM cortex tungkol sa posisyon ng katawan sa espasyo, ay kasangkot sa regulasyon ng paghinga at vascular innervation.
- Bilang karagdagan, ang pagbuo ng reticular ay kinabibilangan ng mga neuron na may mahalagang bahagi sa gawain ng mga sentro ng thermoregulation, kabusugan at kagutuman.

Pangunahing pag-andar
Ang pangunahing layunin ng reticular formation ay sensory analysis ng maraming signal na nagmumula sa iba't ibang bahagi ng GM.
Dahil sa malapit na koneksyon sa spinal cord, ito rin ay tumatagal ng isang aktibong bahagi sa regulasyon ng motor, mula sa swallowing reflex hanggang sa kumplikadong mga operasyon ng motor. Bilang karagdagan, ang reticular formation ay nagsasagawa ng isang activating effect sa buong GM sa kabuuan, na nakikilahok sa regulasyon ng mga cycle ng pagtulog at paggising.
Sa pangkalahatan, ang mga pag-andar ng reticular formation ay ang mga sumusunod:
- Regulasyon ng mga kalamnan ng kalansay (kasangkot sa pagkontrol sa mga galaw ng katawan) at mga autonomic na function (paghinga, pagbahin, sirkulasyon ng dugo, atbp.).
- Pagkontrol sa mga proseso ng pagtulog at pagpupuyat (dahil sa pagkakaloob ng isang pag-activate at pagbabawal na epekto sa cerebral cortex).
- Ang pag-activate ng function (na ipinakita sa katotohanan na ang reticular formation ay nagbibigay ng patuloy na tonic excitation ng GM cortex, dahil kung saan posible na mapanatili ang atensyon, kamalayan at ang kurso ng mga proseso ng pag-iisip.)
- Pagproseso ng mga signal mula sa panlabas at panloob na kapaligiran.
Ang mga natatanging tampok ng gawain ng pagbuo ng reticular ay nauugnay, una sa lahat, kasama ang ilang mga katangian nito:
Ang mga nerve cell na bumubuo sa reticular formation ay may mas mataas na kapasidad para sa tonic stimulation. Nangangahulugan ito na ang karamihan sa mga neuron ay nasa patuloy na kaguluhan at bumubuo ng mga nerve impulses na ipinapadala sa mga nakapatong na bahagi ng GM. Ang tonic na aktibidad na ito ay dahil sa maraming mga kadahilanan:
- Ang pagkamatagusin ng isang malaking bilang ng mga signal sa pamamagitan ng mga istruktura ng reticular formation. Narito ang isang simpleng pagkakatulad: Isipin ang isang grand piano o iba pang instrumentong may kwerdas. Malinaw na kapag direktang hinawakan natin ang mga kuwerdas, nagsisimula silang mag-vibrate at gumawa ng tunog. Ang parehong bagay ay nangyayari sa mga selula ng nerbiyos kapag ang mga senyales mula sa ibang mga neuron ay dumating sa kanila. Gayunpaman, isipin pa natin na hindi natin direktang hinahawakan ang mga string ng instrumento, ngunit, sabihin nating, tumatalon sa tabi nito, malakas na ibinabagsak ang ating mga paa sa sahig. . Maaaring hindi natin marinig ang tunog ng instrumento, ngunit isang bahagya pa ring kapansin-pansing vibration ng mga kuwerdas ang magaganap. Ang parehong bagay ay nangyayari sa mga neuron ng reticular formation. Dahil ang ilang mga signal (parehong afferent at efferent) mula sa iba't ibang mga istraktura ng central nervous system ay patuloy na dumadaan dito, lumilikha ito ng isang pare-parehong tonic excitation ng mga neuron ng reticular formation, dahil sa ang katunayan na ito ay nasa epicenter ng isang pare-parehong neuro - palitan ng salpok.
- Tumaas na sensitivity ng mga neuron sa mga kemikal (mga hormone, gamot, psychotropic substance). Ang isang tasa ng kape na lasing sa umaga ay "i-on" ang mga istruktura ng reticular formation at, dahil sa pangmatagalang pangangalaga ng paggulo sa mga neuron nito, pinapanatili tayong aktibo.
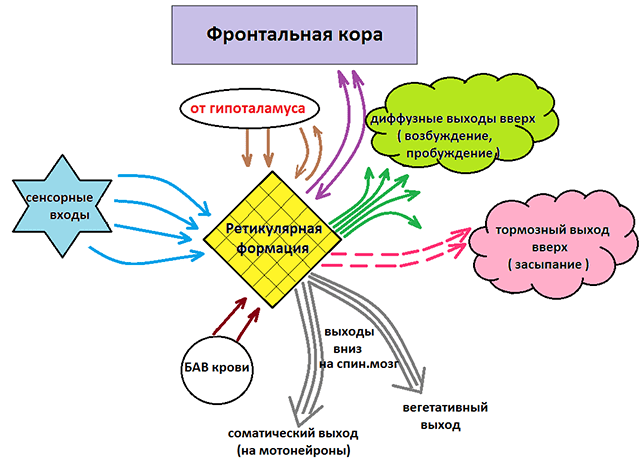
Pababa at pataas na impluwensya ng RF
Tulad ng nabanggit na, ang pagbuo ng reticular ay may kapana-panabik at nagbabawal na epekto sa iba't ibang bahagi ng GM. Sa kasong ito, maaaring makilala ang dalawang departamento na dalubhasa sa paglipat ng paggulo sa ilang mga istruktura ng utak.
Pababang dibisyon: kinakatawan ng mga autonomic at motor center at nagdudulot ng pababang impluwensya sa mga dibisyon ng spinal cord. Ang kaukulang neural accumulations ay kumokontrol sa aktibidad ng respiratory, vasomotor, salivary centers, pati na rin ang mga sentro na responsable para sa pagtatayo ng simple at kumplikadong mga reaksyon ng motor. Ipinapahiwatig nito ang mapagpasyang papel ng gitnang sistema ng nerbiyos sa regulasyon ng kahit elementarya na walang kondisyon na mga reflexes. Ang pagpapasigla ng pababang seksyon ay humahantong sa pagsugpo sa mga sentro ng gulugod at nag-uudyok sa natural na kapaligiran ng isang estado ng malalim na pagtulog (pagtulog "nang walang hulihan binti"). Ang parehong epekto ay maaaring maimpluwensyahan ng artipisyal, halimbawa, sa pamamagitan ng pagpapakilala sa isang tao sa isang estado ng kawalan ng ulirat o kawalan ng pakiramdam.
Pataas na seksyon: kinakatawan ng mga nerve fibers na nagkokonekta sa mga istruktura ng reticular formation sa mga nakapatong na seksyon: ang thalamus, hypothalamus, cerebellum at cortex. Ang pataas na epekto ay may nakapagpapasigla na epekto sa mga istruktura ng cortical at nagbibigay ng aktibong estado ng kamalayan. Ang pataas na impluwensya ay hindi tumitigil kahit na tayo ay natutulog. Kung ang ating utak ay maaaring ganap na "magsara", kung gayon ang bawat paggising ay magiging katulad ng isang kapanganakan: sino ako? Nasaan ako? Paano ako nakarating dito? Gayunpaman, dahil sa gawain ng mga reticular na istruktura, mayroon pa rin tayong pagkakataon na palaging bumalik sa paunang estado ng kamalayan kung saan tayo ay bago ang sandali ng pagtulog. Bilang karagdagan, sa panahon ng pahinga sa gabi, mayroon pa rin tayong kakayahan na tumugon sa ilang mahahalagang stimuli, i.e. kami, bilang isang patakaran, ay hindi natutulog na "patay na pagtulog" at maaaring magising kung ang isang bata ay lumipat at umiyak sa malapit, may nahulog nang malakas, atbp.
Pagpapakita ng pinsala sa mga istruktura
Ang reticular formation ay may mahalagang papel sa integrative na aktibidad ng buong utak. Dahil sa pagganap ng pag-andar ng nangungunang conductor ng lahat ng uri ng nerve impulses sa lahat ng bahagi ng central nervous system, ang reticular formation ay nasa patuloy na operasyon. Ang sobrang mental at emosyonal na labis na karga ay nakakapinsala sa utak sa pangkalahatan at sa reticular formation sa partikular. Sa kabutihang palad, ang napapanahong pangangasiwa ng mga sedative ay maaaring (dahil sa mas mataas na pagkamaramdamin ng mga neuron sa mga epekto ng kemikal) ay mabilis na iwasto ang sitwasyon at gawing normal ang kondisyon.
Gayunpaman, posible rin ang hindi gaanong kanais-nais na mga resulta. Ang pinsala ay posible bilang isang resulta ng craniocerebral trauma, oncological na sakit ng utak, mga nakakahawang sugat.
Ang pangunahing pagpapakita ng kalungkutan ay pagkawala ng kamalayan.
Ang paglabag sa mga pataas na koneksyon ay nagpapakita ng sarili sa isang estado ng kawalang-interes, kahinaan, nadagdagan ang pag-aantok, pagkawatak-watak ng motor, mga kaguluhan sa pagtulog sa gabi. Ang magkakatulad na autonomic disorder ay hindi karaniwan.
Reticular formation - isang hanay ng iba't ibang, na matatagpuan sa kahabaan ng tangkay ng utak, na nagpapagana ng isang pag-activate o pagbabawal na epekto sa iba't ibang mga istruktura ng central nervous system, sa gayon kinokontrol ang kanilang reflex na aktibidad.
Ang reticular formation ng brain stem ay may activating effect sa mga cell at isang inhibitory effect sa motor neurons ng spinal cord. Sa pamamagitan ng pagpapadala ng mga inhibitory at excitatory impulses sa spinal cord sa mga motor neuron nito, ang reticular formation ay nakikilahok sa regulasyon ng skeletal muscle tone.
Ang reticular formation ay nagpapanatili ng tono ng mga autonomic center, nagsasama ng mga sympathetic at parasympathetic na mga impluwensya, naglilipat ng modulating effect mula sa hypothalamus at cerebellum sa mga panloob na organo.
Mga function ng reticular formation
Kontrol ng somatomotor(skeletal muscle activation) ay maaaring direktang sa pamamagitan ng tr. reticulospinalis at indirect through, olives, tubercles ng quadruple, red nucleus, black matter, striatum, thalamic nuclei at kahit somatomotor zones ng cortex.
Somatosensitive control, ibig sabihin. pagbaba sa mga antas ng somatosensory na impormasyon - "mabagal na sakit", pagbabago ng pang-unawa ng iba't ibang uri ng sensory sensitivity (pandinig, paningin, vestibulasyon, amoy).
Kontrol ng visceromotor ang estado ng cardiovascular, respiratory system, ang aktibidad ng makinis na mga kalamnan ng iba't ibang mga panloob na organo.
Neuroendocrine transduction sa pamamagitan ng impluwensya sa mga neurotransmitter, ang mga sentro ng hypothalamus at pagkatapos ay ang pituitary gland.
Mga biorhythms sa pamamagitan ng mga koneksyon sa hypothalamus at pineal gland.
Iba-iba functional na estado ng katawan(pagtulog, paggising, estado ng kamalayan, pag-uugali) ay isinasagawa sa pamamagitan ng maraming koneksyon ng nuclei ng reticular formation sa lahat ng bahagi ng central nervous system.
Koordinasyon gawain ng iba't ibang mga sentro ng brain stem pagbibigay ng mga kumplikadong visceral reflex na tugon (pagbahin, pag-ubo, pagsusuka, paghikab, pagnguya, pagsuso, paglunok, atbp.).
Ang istraktura ng reticular formation
Ang pagbuo ng reticular nabuo sa pamamagitan ng isang hanay ng maraming mga neuron nakahiga nang hiwalay o nakapangkat sa nuclei (tingnan ang Fig. 1 at 2). Ang mga istruktura nito ay naisalokal sa mga gitnang rehiyon ng puno ng kahoy, simula sa itaas na mga segment ng cervical spinal cord hanggang sa itaas na antas ng brainstem, kung saan sila ay unti-unting sumanib sa mga nuclear group. Ang reticular formation ay sumasakop sa mga puwang sa pagitan ng nuclei ng cranial nerves, iba pang nuclei, at mga tract na dumadaan sa brainstem.
Ang mga neuron ng reticular formation ay nailalarawan sa pamamagitan ng isang malawak na iba't ibang mga hugis at sukat, ngunit ang kanilang karaniwang tampok ay ang bumubuo sila ng maraming synaptic contact na may mahabang dendrite at malawak na sumasanga na mga axon, kapwa sa kanilang mga sarili at sa mga neuron sa iba pang nuclei ng utak. Ang mga sangay na ito ay bumubuo ng isang uri ng network ( reticulum), kung saan nagmula ang pangalan - ang reticular formation. Ang mga neuron na bumubuo sa nuclei ng reticular formation ay mayroon mahabang axon na bumubuo ng mga landas patungo sa spinal cord, ang nuclei ng brainstem, at iba pang bahagi ng utak.

kanin. 1. Ang pinakamahalagang structural formations ng midbrain (cross section)
Ang mga neuron ng reticular formation ay tumatanggap ng marami afferent signal mula sa iba't ibang mga istraktura ng central nervous system. Mayroong ilang mga grupo ng mga neuron na tumatanggap ng mga signal na ito. ito pangkat ng mga neuron ng lateral nucleus reticular formation, na matatagpuan sa medulla oblongata. Ang mga nucleus neuron ay tumatanggap ng mga afferent signal mula sa mga interneuron ng spinal cord at bahagi ng isa sa mga hindi direktang spinocerebellar pathway. Bilang karagdagan, nakakatanggap sila ng mga signal mula sa vestibular nuclei at maaaring isama ang impormasyon tungkol sa estado ng aktibidad ng mga interneuron na nauugnay sa mga motor neuron ng spinal cord, at tungkol sa posisyon ng katawan at ulo sa espasyo.
Ang susunod na grupo ay mga neuron ng reticulotegmental nucleus matatagpuan sa hangganan ng dorsal edge ng tulay. Nakatanggap sila ng mga afferent synaptic input mula sa mga neuron ng pretectal nuclei at ang superior hillocks ng quadruple at ipinapadala ang kanilang mga axon sa mga istruktura ng cerebellum na kasangkot sa kontrol ng paggalaw ng mata.
Mga neuron ng reticular formation makatanggap ng iba't ibang signal sa pamamagitan ng mga pathway na nagkokonekta sa kanila sa cerebral cortex (corticoreticulospinal pathways), substantia nigra, atbp.

kanin. 2. Ang lokasyon ng ilang nuclei sa stem ng utak at hypothalamus: 1 - paraventricular; 2 - dorsomedial: 3 - preoptic; 4 - supraoptic; 5 - likod
Bilang karagdagan sa mga inilarawan na afferent pathway, ang mga signal ay ipinapadala sa reticular formation sa pamamagitan ng axonal collaterals mga landas ng sensory system. Kasabay nito, ang mga signal mula sa iba't ibang mga receptor (tactile, visual, auditory, vestibular, sakit, temperatura, proprioceptors, receptors ng mga panloob na organo) ay maaaring magtagpo sa parehong isa.
Mula sa listahan sa itaas ng mga pangunahing afferent na koneksyon ng reticular formation sa iba pang mga lugar ng central nervous system, makikita na ang estado ng tonic neuronal na aktibidad nito ay natutukoy sa pamamagitan ng pag-agos ng halos lahat ng mga uri ng sensory signal mula sa sensory neurons, pati na rin ang mga senyales mula sa karamihan ng mga istruktura ng central nervous system.
Pag-uuri ng reticular formation depende sa direksyon ng mga hibla|
Mga kagawaran |
Katangian |
|
Pababang departamento |
Mga sentro ng halaman:
Mga sentro ng motor:
|
|
Pataas na departamento |
Reticulothalamic Reticulohypothalamic Reticulocerebellar Reticulocortical: pag-activate; hypnogenic |
Ang nuclei ng reticular formation at ang kanilang mga function
Sa loob ng mahabang panahon ay pinaniniwalaan na ang reticular formation, ang istraktura na kung saan ay nailalarawan sa pamamagitan ng malawak na interneuronal na koneksyon, ay nagsasama ng mga signal ng iba't ibang mga modalidad nang hindi naglalabas ng tiyak na impormasyon. Gayunpaman, nagiging mas at mas halata na ang pagbuo ng reticular ay hindi lamang morphologically, ngunit din functionally heterogenous, bagaman ang mga pagkakaiba sa pagitan ng mga pag-andar ng mga indibidwal na bahagi nito ay hindi kasing halata tulad ng karaniwan para sa iba pang mga lugar ng utak.
Sa katunayan, maraming mga neuronal na grupo ng reticular formation ang bumubuo sa nuclei nito (mga sentro) na gumaganap ng mga partikular na function. Ito ang mga neural group na bumubuo sentro ng vasomotor medulla oblongata (higanteng cell, paramedian, lateral, ventral, caudal nuclei ng medulla oblongata), sentro ng paghinga(higanteng cell, maliit na cell nucleus ng medulla oblongata, oral at caudal nuclei ng pons), mga sentro ng pagnguya at paglunok(lateral, paramedian nucleus ng medulla oblongata), mga sentro ng paggalaw ng mata(paramedian na bahagi ng tulay, rostral na bahagi ng midbrain), mga sentro ng regulasyon ng tono ng kalamnan(ang rostral nucleus ng pons at ang caudal nucleus ng medulla oblongata), atbp.
Ang isa sa pinakamahalagang di-tiyak na pag-andar ng reticular formation ay regulasyon ng pangkalahatang aktibidad ng neural ng cortex at iba pang mga istruktura ng central nervous system. Sa pagbuo ng reticular, ang biological na kahalagahan ng mga papasok na sensory signal ay tinasa, at, depende sa mga resulta ng pagtatasa na ito, maaari itong buhayin o pagbawalan, sa pamamagitan ng hindi tiyak o tiyak na mga neuronal na grupo ng thalamus, mga proseso ng neural sa buong cerebral cortex o sa lahat ng mga indibidwal na zone. Samakatuwid, ang stem reticular formation ay tinatawag din sistema ng pag-activate ng bariles utak. Dahil sa mga katangiang ito, ang pagbuo ng reticular ay maaaring maka-impluwensya sa antas ng pangkalahatang aktibidad ng cortex, ang pagpapanatili nito ay ang pinakamahalagang kondisyon para sa pagpapanatili ng kamalayan, ang estado ng pagkagising, at ang pagbuo ng pokus ng pansin.
Ang pagtaas sa aktibidad ng reticular formation (laban sa isang pangkalahatang mataas na background) sa mga indibidwal na sensory, nag-uugnay na mga lugar ng cortex ay nagbibigay ng kakayahang ihiwalay at iproseso ang partikular na impormasyon na pinakamahalaga sa isang naibigay na oras para sa katawan at ayusin ang sapat na pag-uugali ng pagtugon mga reaksyon. Karaniwan, ang mga reaksyong ito, na inayos kasama ang paglahok ng reticular formation ng brainstem, ay nauuna sa orientational na paggalaw ng mga mata, ulo at katawan sa direksyon ng pinagmulan ng signal, mga pagbabago sa paghinga at sirkulasyon ng dugo.
Ang pag-activate ng epekto ng reticular formation sa cortex at iba pang mga istruktura ng central nervous system ay isinasagawa kasama ang mga pataas na landas na nagmumula sa higanteng cell, lateral at ventral reticular nuclei ng medulla oblongata, pati na rin mula sa nuclei ng pons. at midbrain. Kasama ang mga landas na ito, ang mga stream ng nerve impulses ay isinasagawa sa mga neuron ng nonspecific na nuclei ng thalamus at, pagkatapos ng kanilang pagproseso, ay inililipat sa thalamic nuclei para sa kasunod na paghahatid sa cortex. Bilang karagdagan, mula sa nakalistang reticular nuclei, ang mga daloy ng signal ay isinasagawa sa mga neuron ng posterior hypothalamus at basal ganglia.
Bilang karagdagan sa pag-regulate ng aktibidad ng neural ng mas matataas na bahagi ng utak, ang reticular formation ay maaaring umayos ng mga sensory function. Ginagawa ito sa pamamagitan ng pag-impluwensya sa pagpapadaloy ng mga afferent signal sa mga nerve center, ang excitability ng mga neuron ng mga nerve center, pati na rin ang sensitivity ng mga receptor. Ang pagtaas sa aktibidad ng reticular formation ay sinamahan ng isang pagtaas sa aktibidad ng mga neuron ng sympathetic nervous system, na nagpapasigla sa mga organo ng pandama. Bilang resulta, maaaring tumaas ang visual acuity, pandinig, tactile sensitivity.
Kasama ang pataas na pag-activate at pag-iwas sa mga impluwensya sa mas mataas na bahagi ng utak, ang reticular formation ay nakikibahagi sa regulasyon ng mga paggalaw, na nagbibigay ng mga epekto sa pag-activate at pagbabawal sa spinal cord. Sa nuclei nito, ang parehong mga pataas na landas mula sa proprioceptors at spinal cord patungo sa utak at ang pababang mga daanan ng motor mula sa cerebral cortex, basal nuclei, cerebellum at pulang nucleus ay inililipat. Bagaman ang mga pataas na neural pathway na humahantong mula sa reticular formation hanggang sa thalamus at cortex ay pangunahing gumaganap sa pagpapanatili ng pangkalahatang antas ng aktibidad ng cerebral hemispheres ng utak, ngunit ito ang function ng mga ito na mahalaga para sa nakakagising cortex upang magplano. , simulan, isagawa ang mga paggalaw at kontrolin ang kanilang pagpapatupad. Mayroong isang malaking bilang ng mga collateral na koneksyon sa pagitan ng pataas at pababang mga landas sa pamamagitan ng reticular formation, kung saan maaari silang magkaroon ng impluwensya sa isa't isa. Ang pagkakaroon ng ganoong malapit na pakikipag-ugnayan ay lumilikha ng mga kondisyon para sa magkaparehong impluwensya ng rehiyon ng reticular formation, na nakakaimpluwensya sa pamamagitan ng thalamus sa aktibidad ng cortex, na nagpaplano at nagpapasimula ng mga paggalaw, at ang rehiyon ng reticular formation, na nakakaimpluwensya sa executive neural na mekanismo ng spinal cord. Sa reticular formation mayroong mga grupo ng mga neuron na nagpapadala ng karamihan sa mga axon sa cerebellum, na kasangkot sa regulasyon at koordinasyon ng mga kumplikadong paggalaw.
Kasama ang pababang reticulospinal pathways, ang reticular formation ay direktang nakakaapekto sa mga function ng spinal cord. Ang direktang impluwensya sa mga sentro ng motor nito ay isinasagawa ng medial reticulospinal tract, nagmumula sa nuclei ng pons at pangunahing pinapagana ang inter- at y-motor neuron ng mga extensor at pinipigilan ang mga motor neuron ng flexor na kalamnan ng trunk at limbs. Sa pamamagitan ng lateral reticulospinal tract simula sa giant cell nucleus ng medulla oblongata, ang reticular formation ay may activating effect sa inter- at y-motor neurons ng flexor muscles ng limbs at pinipigilan ang neurons ng extensor muscles.
Mula sa mga eksperimentong obserbasyon sa mga hayop, alam na ang pagpapasigla ng mas maraming rostrally na matatagpuan na mga neuron ng reticular formation sa antas ng medulla oblongata at midbrain ay may diffuse facilitating effect sa spinal reflexes, at stimulation ng mga neuron sa caudal na bahagi ng medulla oblongata ay sinamahan ng pagsugpo ng synal reflexes.
Ang pag-activate at pagbabawal na epekto ng reticular formation sa mga motor center ng spinal cord ay maaaring isagawa sa pamamagitan ng y-motor neurons. Sa kasong ito, ang mga reticular neuron ng rostral na rehiyon ng reticular formation ay nagpapagana ng mga y-motor neuron, na nagpapasigla sa intrafusal. mga hibla ng kalamnan, maging sanhi ng kanilang pag-urong, buhayin ang mga receptor ng spindle ng kalamnan. Ang daloy ng mga signal mula sa mga receptor na ito ay nagpapagana ng mga a-motoneuron at nagiging sanhi ng pagkontrata ng kaukulang kalamnan. Ang mga neuron sa caudal region ng reticular formation ay pumipigil sa aktibidad ng y-motor neurons ng spinal cord at nagiging sanhi ng relaxation ng kalamnan. Ang pamamahagi ng tono sa malalaking grupo ng kalamnan ay nakasalalay sa balanse ng aktibidad ng neuronal sa mga lugar na ito ng pagbuo ng reticular. Dahil ang balanse na ito ay nakasalalay sa mga pababang impluwensya sa reticular formation ng cerebral cortex, basal ganglia, hypothalamus, cerebellum, ang mga istruktura ng utak na ito ay maaari ding makaapekto sa pamamahagi ng tono ng kalamnan at postura ng katawan sa pamamagitan ng reticular formation at iba pang nuclei ng stem ng utak.
Ang malawak na sumasanga ng mga axon ng mga reticulospinal pathway sa spinal cord ay lumilikha ng mga kondisyon para sa impluwensya ng reticular formation sa halos lahat ng mga neuron ng motor at, nang naaayon, sa estado ng mga kalamnan ng iba't ibang bahagi ng katawan. Ang tampok na ito ay nagbibigay ng isang epektibong epekto ng reticular formation sa reflex distribution ng tono ng kalamnan, pustura, oryentasyon ng ulo at katawan sa direksyon ng panlabas na stimuli at ang pakikilahok ng reticular formation sa pagpapatupad ng mga boluntaryong paggalaw ng kalamnan sa mga proximal na bahagi. ng katawan.
Sa gitnang bahagi ng reticular giant cell nucleus, mayroong isang site, ang pangangati na pumipigil sa lahat ng mga motor reflexes ng spinal cord. Ang pagkakaroon ng naturang pagsugpo sa mga istruktura ng utak sa spinal cord ay natuklasan ni I.M. Sechenov sa mga eksperimento sa mga palaka. Ang kakanyahan ng mga eksperimento ay pag-aralan ang estado ng spinal cord reflexes pagkatapos ng transection ng brain stem sa antas ng diencephalon at pangangati ng caudal section ng incision na may kristal ng sodium chloride. Ito ay lumabas na ang motor spinal reflexes ay hindi lumitaw sa panahon ng pagpapasigla o naging weakened at nakuhang muli pagkatapos ng pag-aalis ng pangangati. Kaya, ito ay ipinahayag sa unang pagkakataon na ang isang nerve center ay maaaring pagbawalan ang aktibidad ng isa pa. Pinangalanan ang phenomenon na ito gitnang pagpepreno.
Ang reticular formation ay gumaganap ng isang mahalagang papel sa regulasyon ng hindi lamang somatic, kundi pati na rin ang mga autonomic function (ang reticular nuclei ng stem ng utak ay bahagi ng istraktura ng mga mahahalagang bahagi ng respiratory center at mga sentro ng regulasyon ng sirkulasyon ng dugo). Ang lateral group ng pons reticular nuclei at ang dorsolateral tegmental nucleus ay nabuo sentro ng ihi ng tulay. Ang mga axon ng mga neuron ng nuclei ng sentrong ito ay umaabot sa preganglionic neuron ng sacral spinal cord. Ang pagpapasigla ng mga neuron ng mga nuclei na ito sa pons ay sinamahan ng isang pag-urong ng kalamnan ng dingding ng pantog at pag-ihi.
Sa dorsolateral bridge, matatagpuan ang parabrachial nucleus, sa mga neuron kung saan nagtatapos ang mga fibers ng sensory neurons ng panlasa. Ang mga neuron ng nucleus, tulad ng mga neuron ng bluish spot at ang substantia nigra, ay naglalaman ng neuromelanin. Ang bilang ng mga naturang neuron sa parabrachial nucleus ay bumababa sa Parkinson's disease. Ang mga neuron ng parabrachial nucleus ay may mga koneksyon sa mga neuron ng hypothalamus, amygdala, suture nuclei, solitary tract at iba pang nuclei ng brain stem. Ipinapalagay na ang parabrachial nuclei ay nauugnay sa regulasyon ng mga autonomic function at ang pagbaba sa kanilang bilang sa parkinsonism ay nagpapaliwanag ng paglitaw ng mga autonomic disorder sa sakit na ito.
Ipinakita sa mga eksperimento sa mga hayop na ang pagpapasigla ng ilang mga lokal na lugar ng mga reticular na istruktura ng medulla oblongata at ang pons ay maaaring maging sanhi ng pagsugpo sa aktibidad ng cortex at pagtulog. Sa kasong ito, lumilitaw ang mga low-frequency (1-4 Hz) wave sa EEG. Sa batayan ng inilarawan na mga katotohanan, pinaniniwalaan na ang pinakamahalagang pag-andar ng pataas na impluwensya ng reticular formation ay ang regulasyon ng cycle ng sleep-wakefulness at ang antas ng kamalayan. Ito ay lumabas na ang isang bilang ng mga nuclei ng reticular formation ng stem ng utak ay direktang nauugnay sa pagbuo ng mga kondisyong ito.
Kaya, sa bawat panig ng gitnang tahi ng tulay, mayroong paramedian reticular nuclei, o suture core naglalaman ng mga serotonergic neuron. Sa caudal na bahagi ng tulay, kasama nila ang lower central nucleus, na isang pagpapatuloy ng nucleus ng suture ng medulla oblongata, at sa rostral na bahagi ng tulay, ang upper central nucleus, na tinatawag na ankylosing spondylitis, o ang median nucleus ng tahi, ay bahagi ng nuclei ng pons suture.
Sa rostral na bahagi ng pons, sa dorsal na bahagi ng operculum, mayroong isang pangkat ng mga nuclei maasul na lugar. Naglalaman ang mga ito ng humigit-kumulang 16,000-18,000 melanin-containing noradrenergic neurons, ang mga axon na kung saan ay malawak na kinakatawan sa iba't ibang bahagi ng central nervous system - ang hypothalamus, hippocampus, cerebral cortex, cerebellum at spinal cord. Ang mala-bughaw na lugar ay umaabot sa midbrain, at ang mga neuron nito ay sinusubaybayan sa espasyo ng midbrain. Ang bilang ng mga neuron sa nucleus ng bluish spot ay bumababa sa parkinsonism, Alzheimer's disease at Down's syndrome.
Parehong serotonergic at norepinephrine neuron sa reticular formation ay may papel sa pagkontrol sa sleep-wake cycle. Ang pagsugpo sa synthesis ng serotonin sa nuclei ng seam ay humahantong sa pag-unlad ng hindi pagkakatulog. Ipinapalagay na ang mga serotonergic neuron ay bahagi ng neural network na kumokontrol sa slow-wave sleep. Sa pagkilos ng serotonin sa mga neuron ng mala-bughaw na lugar, nangyayari ang kabalintunaan ng pagtulog. Ang pagkasira ng nuclei ng mala-bughaw na lugar sa mga eksperimentong hayop ay hindi humahantong sa pag-unlad ng hindi pagkakatulog, ngunit nagiging sanhi ng pagkawala ng paradoxical na yugto ng pagtulog sa loob ng ilang linggo.
Ang network ng mga nerve cells na matatagpuan sa gitna ng brain stem ay inilarawan ng Hungarian anatomist na si Jozsef Lenhossek noong 1855. Phylogenetically, ito ay isang sinaunang grupo ng mga cell na mayroon ang lahat ng vertebrates at gumaganap ng mahalagang papel sa regulasyon ng aktibidad ng central nervous sistema,
Sa gitna ay may mga reticular cells; sa mga gilid - pandama na landas. Sa mga tao, ang reticular formation ay nagsisimula sa itaas ng spinal cord sa medulla oblongata at umaabot sa diencephalon sa pamamagitan ng tulay at midbrain. Ang kakaiba ng pormasyon na ito ay na sa gilid (mula sa mga gilid) ay napapalibutan ito ng mga pandama na landas na inilarawan "sa unang bahagi ng aklat. Kaya, ang mga hibla ng reticular formation ay nasa gitna at napapalibutan sa labas ng isang layer ng sensory. Ang neural network ng reticular formation ay napaka heterogenous: ang laki ng mga neuron ay nag-iiba mula 10 hanggang 100 microns, at sila ay matatagpuan din sa iba't ibang paraan. Sa reticular formation, ang parehong mabilis at mabagal na pagpapadaloy ng mga impulses ay posible. opinyon ng karamihan sa mga may-akda, ito ay wala ng anumang organisasyon: sa nagkakalat na pinahabang pormasyon na ito, walang tiyak na mga istruktura ang maaaring makilala. Kamakailan lamang, gayunpaman, iminungkahi ng mga mananaliksik ang pagkakaroon ng magkakahiwalay na grupo ng mga selula sa reticular formation. Gayunpaman, lahat ay sumasang-ayon pa rin na ang mga pagkakaiba ay gumagana lamang: sa ilang mga lugar ay matatagpuan ang inhibitory nuclei, at sa iba pa - activating nuclei.
Maramihang koneksyon. Si Cahal ang unang nakarating sa konklusyon na ang lahat ng sensory pathways mula sa spinal cord at medulla oblongata hanggang sa cortex ay nagbibigay ng mga collateral sa activating reticular system. Para sa reticular activation, ang mga collateral na ito ay pinakamahalaga. Ang mga collateral sa reticular formation ay napupunta rin mula sa cortex, thalamus at cerebellum. Sa bahagi nito, ang reticular formation ay nagpapadala rin ng mga hibla sa cortex, gayundin sa mga motor neuron ng spinal cord.
"Mga gising na neuron". Ang pag-andar ng pag-activate ng reticular formation ay unang ipinakita sa mga eksperimento ng Maguna at Moruzzi noong 1949. Pinasigla nila ang reticular system sa pamamagitan ng mga electrodes na itinanim sa stem ng utak, at sa gayon ay nagdulot ng isang tugon sa paggising, iyon ay, nagising ang isang natutulog na hayop. Kasabay nito, ang paglipat mula sa alpha hanggang beta na aktibidad ay maaaring maobserbahan sa larawan ng EEG. Ang pag-uugali ng hayop ay nagbago nang naaayon.
Ascending activating system. Kapansin-pansin, ang paggising ay maaaring ma-trigger ng nakahiwalay na pagpapasigla ng reticular system kahit na pinutol ang mga sensory pathway. Samantala, sa kabaligtaran na kaso - pagkatapos ng pinsala sa reticular formation ng isang high-frequency na electric current, na buo ang lahat ng sensory pathways - walang stimulation ang gumising sa hayop mula sa malalim na pagtulog. Ang pagtuklas ng Megun at Moruzzi ay natagpuan na may kaugnayan sa obserbasyon ni Bremer na inilarawan sa itaas at sa Pavlovian orientation reflex. Ang sistema sa stem ng utak, na sumusuporta
ang tono ng cerebral cortex, tinawag ni Magun ang ascending activating system (Fig. 37).
Paggising, ngunit hindi impormasyon. Ang antas ng aktibidad ng reticular ay natutukoy ng eksklusibo sa pamamagitan ng mga impulses na pumapasok sa reticular formation kasama ang mga collateral ng mga sensory pathway, ibig sabihin, panlabas na stimuli. Ang reticular formation ay wala sa sarili nitong kusang aktibidad. Ang tungkulin nito ay "gisingin" ang buong masa ng mga neuron sa parehong cerebral hemispheres. Ang pagpupuyat ay,
Somatosensory
balat ^ Thalamus
marahil ay isang function ng cortex; ang reticular formation ay "kabilang" lamang ng mga cortical neuron. Ito ay isang pangkalahatang "alarm system" na tumutugon sa lahat ng sensory impulses sa parehong paraan, hindi alintana kung ang mga ito ay nagmula sa auditory, visual o skin receptors. Ang tungkulin nito ay gisingin ang cortex, hindi upang ihatid ang tiyak na impormasyon.
Mag-trigger ng alarm reaction. Malamang na ang pag-activate ng reticular formation ay nauugnay din sa mga mekanismo ng endocrine. Halimbawa, pagkatapos ng pagpapakilala ng adrenaline sa mga eksperimentong hayop, ang reticular activation ay maaaring hatulan ng EEG desynchronization. Ipinakita na ang adrenaline, iyon ay, nadagdagan na aktibidad ng nagkakasundo, ay palaging nagiging sanhi ng pag-activate ng reticular, na nagpapahusay sa estado ng paggising ng hayop. Tinawag ng Cannon ang nagresultang kumplikadong pagbabago ng mga function ng katawan bilang isang alarma na reaksyon. Ang kundisyong ito ay nagpapakita mismo sa acceleration
rate ng puso, pagtaas ng presyon ng dugo at temperatura ng katawan, hyperglycemia, atbp. Kaya, ang pag-activate ng reticular system ay maaaring magdulot ng pagtaas sa tono ng nagkakasundo, na kung saan ay pinahuhusay ang estado ng pagkagising at ang pagiging handa ng buong organismo para sa proteksyon.
Proteksyon mula sa mga kadahilanan ng stress. Dahil sa trabaho ni Selye, ito ay naging kilala na ang buong hormonal system - lalo na ang pituitary gland, na matatagpuan sa base ng bungo, at ang adrenal cortex - ay isinaaktibo ng isang chain reaction, na kung saan ay na-trigger ng mas mataas na nagkakasundo aktibidad. Samakatuwid, malamang na ang buong kumplikadong sistema ng pagtatanggol at pagbagay ay kinokontrol ng pag-activate ng reticular formation. Ang mapaminsalang panlabas na stimuli (mga epekto ng stress) ay nagpapagana sa pagbuo ng reticular, at sa gayon din ang cortex, kasama ang mga collateral ng mga sensory pathway. Kasabay nito, tumataas ang aktibidad ng nagkakasundo at inilabas ang adrenaline, na nagpapahaba sa estado ng gising ng cortex. Kaya, ang nonspecific hormonal defense mechanism na inilarawan ni Cannon at Selye at ang reticular activating system ay functionally interrelated.
Kontrol ng posisyon ng katawan. Ang reticular activating system ay hindi lamang kasangkot sa hormonal regulation, ngunit gumaganap din ng iba pang mga function. Ang pakikilahok nito sa kontrol ng aktibidad ng motor at pagpapanatili ng tono ng kalamnan ay pinag-aralan nang detalyado. Ang mga rehiyon ng cortex at subcortical ay naglalaman ng isang bilang ng mga espesyal na sentro ng motor na bahagi ng isang kumplikadong sistema ng motor na nagre-regulate sa sarili na kinabibilangan ng maraming bahagi mula sa spinal cord hanggang sa cortex. Ang cerebellum ay gumaganap din ng isang mahalagang papel sa sistemang ito. Ang pinakamababa - spinal - ang antas ng efferent motor system ay nasa ilalim ng kontrol ng pag-activate ng reticular formation. Ang kontrol na ito ay isinasagawa ng dalawang uri ng mga impluwensya.
Una, ang reticular formation ay nakakaapekto sa mga reflexes na kumokontrol sa posisyon ng katawan sa kabila ng gravity at iba pang mga deflecting pwersa (nabanggit na namin ang mga reflexes na ito na may kaugnayan sa proprioceptors na bumubuo ng afferent link ng cortical control ng aktibidad ng motor). Ang kontrol na ito ay malamang na isinasagawa ng gamma-efferent fibers na papunta sa mga spindle ng kalamnan. Pangalawa, ang mga impluwensya ng reticular ay may mahalagang papel sa pagpapanatili ng tono ng kalamnan sa pahinga. Ang mga kalamnan ng kalansay ay hindi nakakarelaks, kahit na sila ay hindi aktibo, ngunit nasa isang estado ng ilang pangkalahatang tono. Maihahambing ang mga ito sa mga kuwerdas ng isang grand piano, na nakaunat bago pa man magawa ang anumang tunog. Ang afferent na aktibidad ng proprioceptors (muscle spindles) ay gumaganap ng napakahalagang papel sa regulasyon ng tono ng kalamnan (tingnan ang p. 57). Kaya
Kaya, sa mga nagtatanggol na reaksyon o sa pagsalakay, ang reticular formation ay hindi lamang nagpapagana sa cortex at nagpapalitaw ng mga reaksyon ng hormonal, ngunit inilalagay din ang mga kalamnan ng kalansay sa isang estado ng pagiging handa.
Regulasyon ng aktibidad ng autonomic nervous system. Ang mga pag-andar na inilarawan sa itaas ay nagpapahiwatig ng kahalagahan ng medyo maliit na grupong ito ng mga neuron. Ngunit, bilang karagdagan, ang mga sentro ng mahahalagang function na matatagpuan sa medulla oblongata ay konektado sa reticular formation: mga neuron na kumokontrol sa paghinga at nuclei na kumokontrol sa sirkulasyon ng dugo at iba't ibang mga mekanismo ng pagtunaw. Kaya, masasabi na ang diffuse reticular system ay nagsasama ng lahat ng autonomic na aktibidad na kinokontrol ng stem ng utak.
Mga epekto sa pagpepreno. Tulad ng nabanggit na, ang mga stimuli na nag-uudyok sa aktibidad ng salpok sa extero- at interoceptors ay nagpapagana sa pagbuo ng reticular, na kung saan ay pinahuhusay ang aktibidad ng cortex. Gayunpaman, ang ilang mga afferent impulses, sa kabaligtaran, ay binabawasan ang aktibidad ng reticular at sa gayon ay binabawasan ang tono ng cortex. Ang pagsusuri ng mga fibers IX at X ng cranial nerves ay nagsiwalat ng mga sensory fibers na nagmumula sa dingding ng aorta at carotid artery at ang pagpapasigla nito ay humahantong sa isang pagpapahina ng aktibidad ng reticular.
Ang physiological function ng mga inhibitory effect na ito ay hindi lubos na malinaw. Marahil sila ay bahagi ng inhibitory apparatus (Moruzzi et al.), Na responsable para sa estado ng pagtulog. Ang mga inhibitory reticular neuron ay matatagpuan sa loob at labas ng reticular formation. Ang isyung ito ay tatalakayin nang detalyado sa kabanata sa pagtulog. Dito, ang mga mekanismo ng pagbabawal ay binanggit lamang bilang bahagi ng self-regulating apparatus ng reticular formation.
Regulasyon sa sarili. Ang organismo ng isang hayop o isang tao, dahil sa kakayahang mag-regulasyon sa sarili, ay maaaring ituring bilang isang awtomatikong sistema. Sa ganoong sistema, ang regulasyon ay isang proseso kung saan pinapanatili ang katatagan ng ilang mga parameter nito. Ang konsepto ng regulasyon ay lumitaw sa biology bago pa ang panahon ng cybernetics at computer. Bukod dito, ang iba't ibang mga biological system ay ginamit bilang mga modelo para sa disenyo ng mga awtomatikong regulator. Ang prinsipyo ng homeostasis na inilarawan ni Cannon ay ang batayan ng isang bilang ng mga aparato na idinisenyo upang pag-aralan ang self-regulation.
Feedback. Ang pangunahing mekanismo ng self-regulation ay feedback, na nagsisiguro ng tuluy-tuloy na daloy ng impormasyon tungkol sa mga resulta ng mga utos nito sa control center. Ang layunin ng feedback ay upang ayusin ang mga impluwensya sa regulasyon, upang itama ang mga ito ng central regulatory apparatus. Sa teknolohiya, ginagamit ang feedback, sa partikular,
sa mga servomechanism. Imposible ang awtomatikong kontrol nang walang feedback, iyon ay, nang walang control center na tumatanggap ng impormasyon tungkol sa epekto ng mga utos nito. Positibo ang feedback kung humahantong ito sa pagtaas ng resultang paglihis, at negatibo, kung binabawasan nito ang naturang paglihis. Pareho sa mga mekanismo ng feedback na ito ay gumagana sa mga sensory system at sa paggising function.
Serebral Control Center. Ang pinakamahalagang bahagi ng lahat ng self-regulating automatic system ay ang apparatus na kumokontrol sa adaptasyon ng katawan at tinitiyak ang constancy ng mga parameter nito. Ang bahaging ito ay tinatawag na control center. Ang utak ng tao ay maaaring isipin bilang ang hub kung saan binabago ng mga mekanismo ng feedback ang mga utos at sa gayon ay tinitiyak ang pinakamainam na paggana.
Pagpigil sa pagbuo ng reticular. Kung ang pagbuo ng reticular ay itinuturing na isang bloke na kumokontrol sa antas ng pagkagising ng katawan, dapat mayroong isang mekanismo ng feedback na pumipigil sa aktibidad ng paggising nito. Ang ilang mga naturang mekanismo ay inilarawan kamakailan.
Cortical inhibition. Ang una sa mga mekanismong ito ay ang nagbabawal na epekto ng cortex. Sa pagitan ng cortex at ng reticular formation mayroong hindi lamang pataas, kundi pati na rin pababang mga hibla na nagsasagawa ng mga impulses mula sa cortex. Kaya, ang mga reticular impulses na kumokontrol sa tono ng cortex ay kinokontrol mismo ng mga cortical center. Magkasama, ang mga dibisyong ito ay bumubuo ng isang self-regulating system, na binuo sa prinsipyo ng isang reflex ring.
Thalamic inhibition. Ang thalamus ay naglalaman din ng isang nervous network, na para sa kanyang nagkakalat na istraktura ay tinatawag na thalamic reticular formation at kung saan ay kasangkot sa kontrol ng reticular activation. Tulad ng makikita natin, ang mababang-amplitude na high-frequency na EEG na katangian ng wakefulness ay ang resulta ng pagpapasigla ng brainstem reticular formation, at ang high-amplitude na low-frequency na aktibidad sa panahon ng pagtulog ay dahil sa thalamic reticular activity. Ang mga Thalamic reticular cells ay mga antagonist ng reticular formation ng brainstem. Ang elektrikal na pagpapasigla ng mga thalamic neuron na ito ay maaaring magkaroon ng isang nagbabawal na epekto, na, kasama ng iba pang mga epekto, ay humahantong sa mabagal na naka-synchronize na aktibidad ng cortical. Ang mga istrukturang nagbabawal sa Thalamic ay unang inilarawan ng Swiss physiologist na si Hess mga tatlumpung taon na ang nakalilipas.
Medullary inhibition. Ang mga antagonist ng aktibidad ng reticular ay mga inhibitory neuron din sa medulla oblongata, na inilarawan ni Moruzzi. Bagaman ang mga neuron na ito ay bahagi ng reticular formation,

gumaganap sila hindi pag-activate, ngunit pag-andar ng pagbawalan. Ayon kay Moruzzi at iba pa, ang mga medullary inhibitory neuron ay may mahalagang papel sa mekanismo ng pagtulog.
Ang pagpupuyat ng utak ay sinusuportahan ng isang self-regulating system, na binubuo ng reticular formation at ang mga nagbabawal na istruktura na inilarawan sa itaas. Ang antas ng cortical tone, iyon ay, ang kakayahan ng utak na pasiglahin ng sensory input, ay nakasalalay sa balanse sa pagitan ng pag-activate at pagbabawal na mga mekanismo (Larawan 38).



